博文
分子医学基础-不完美的设计-酷蛋白与肿瘤(13-16)
||
13、不完美的设计-酷蛋白与肿瘤
我最早认识ku蛋白时(ku70, ku80/86, “Ku” 是最先发现“Ku"“酷”蛋白患者名字的简称)以为它们是专门被设计成DNA断端识别蛋白,负责与“意外”断裂的DNA断端共价结合,一方面避免外切酶把断裂的DNA降解,另一方面作为DNA-PK激酶的识别信号做出应答(DNA Damage Response, DDR),为利用非同源末端连接机制把断裂的DNA再连接。近年来,包括我们自己的研究发现,事情远非如此简单,除了识别DNA断裂出现断端,以免造成更糟糕的影响外,ku70 参与调节至少199个基因的差异性转录,而ku80,相对较少,即便如此也与至少29个基因的差异性转录调节有关。
而与Ku70-Ku80“联手”参与DNA断端识别和启动NHEJ(non-Homologous End Jioning, NHEJ, 非同源末端连接)的DNA-PK(激酶)也会影响7个基因的差异性转录。
其中,非常值得关注的是,我们发现了Ku70 抑制NOX2 的基因转录,而NOX2作为NOX(NADPH氧化酶)家族成员之一所“制造”的活性氧游离基在“天然免疫”(innate immunity)中发挥重要作用(另一种是利用免疫细胞,免疫因子和抗体的adaptive immunity, 源自一个“转座子”的插入建构的V(D)J重排,见《每个人的医学》化学工业出版社);而另一组中国人组成的研究团队则发现Ku70 的“伙伴”Ku80则与COX2(前列腺素环氧化合成酶)的基因转录有关(COX2本身负责前列腺素合成,受炎性因子有关的白介素等的调节,而前列腺素PGE2不止与炎症情况下的红肿热痛等等有关,而且它还参与细胞繁殖、凋亡,肿瘤细胞的浸润等过程)。
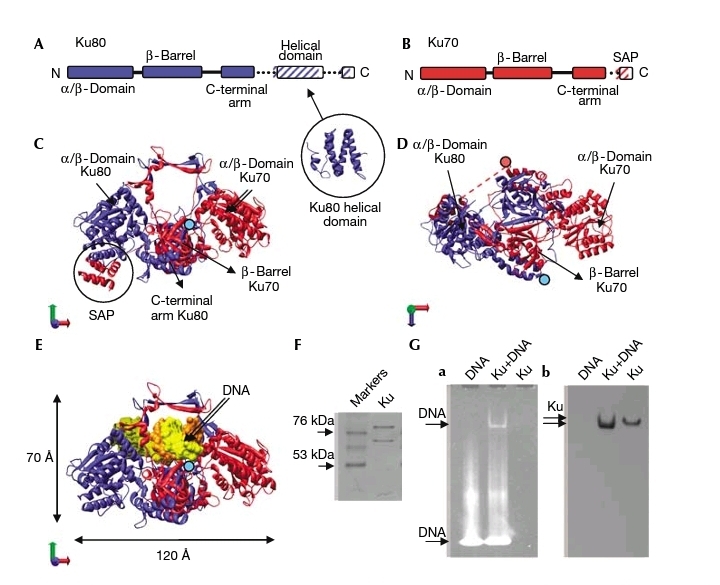 Ku70 ku80的分子结构及与DNA结合方式
Ku70 ku80的分子结构及与DNA结合方式
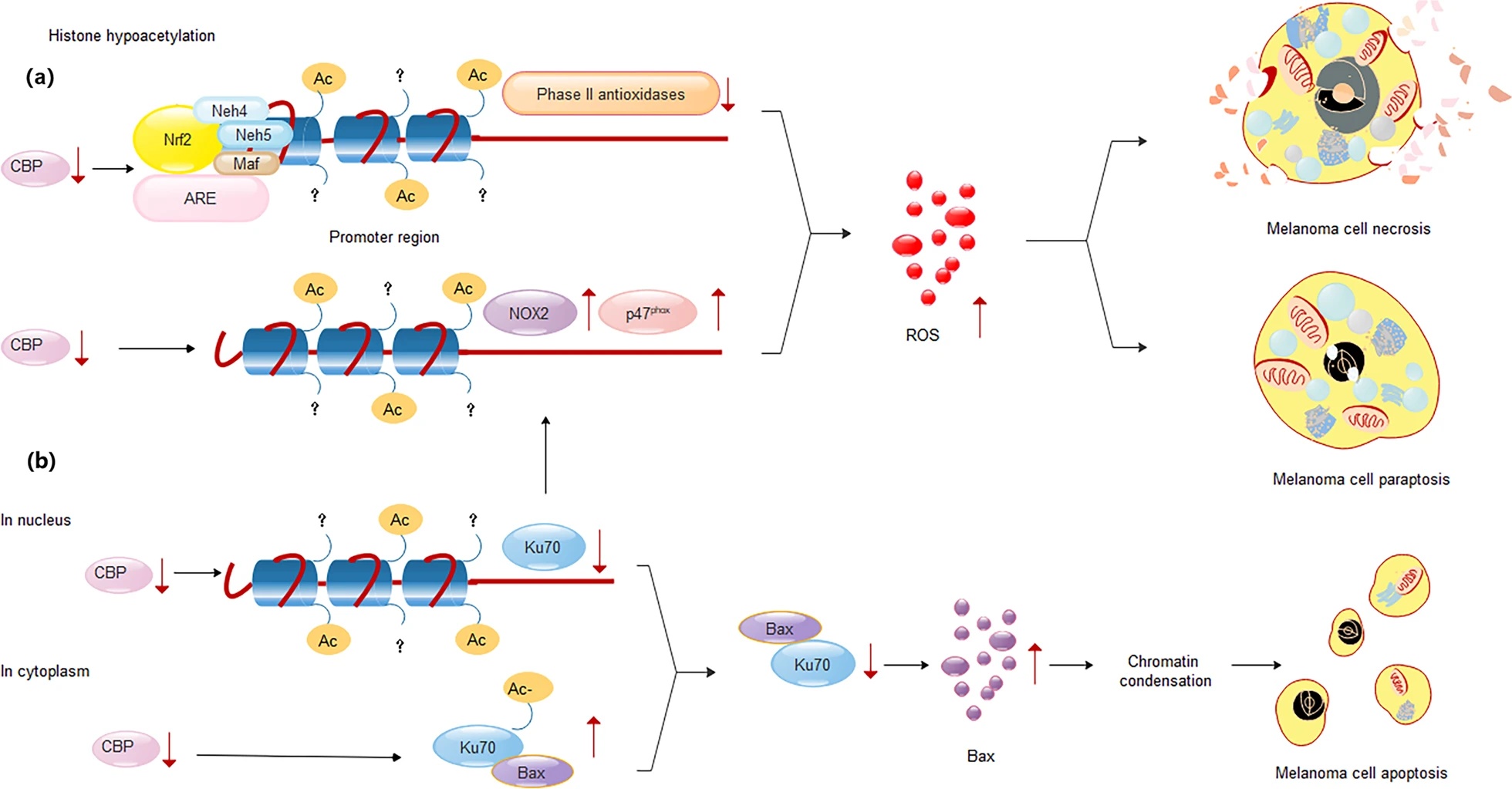 我们发现Ku70-CBP(一类广泛的蛋白乙酰基转移酶)影响nox2基因的转录避免坏死、副凋亡;通过Ku70与BAX结合成复合体阻止其启动线粒体凋亡
我们发现Ku70-CBP(一类广泛的蛋白乙酰基转移酶)影响nox2基因的转录避免坏死、副凋亡;通过Ku70与BAX结合成复合体阻止其启动线粒体凋亡
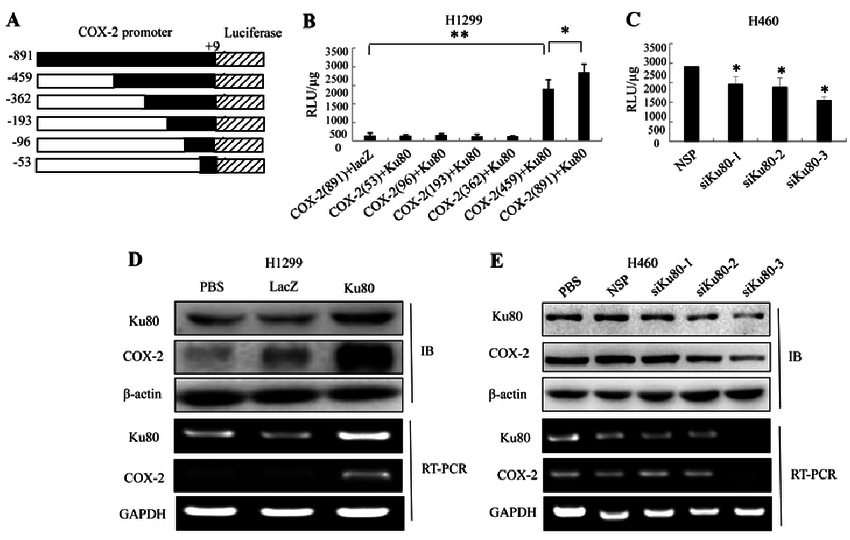 Ku80促进COX2转录 大连医科大学第一附院
Ku80促进COX2转录 大连医科大学第一附院
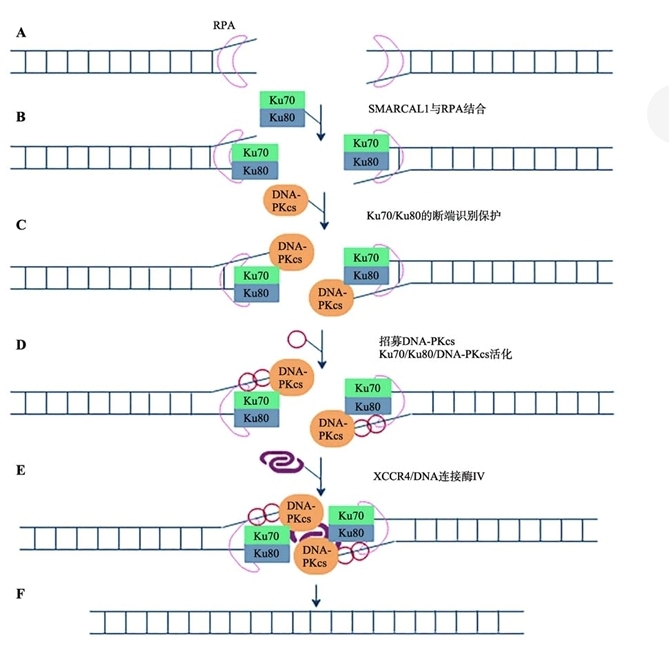 Ku70-Ku80负责识别DNA断端
Ku70-Ku80负责识别DNA断端
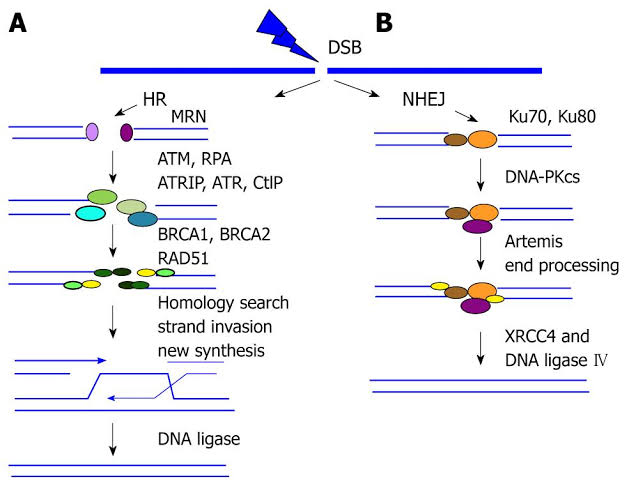 DNA断端有些被MRN识别有些被Ku70 -K80识别当前没有确切的数据表征两类断端的识别特征
DNA断端有些被MRN识别有些被Ku70 -K80识别当前没有确切的数据表征两类断端的识别特征
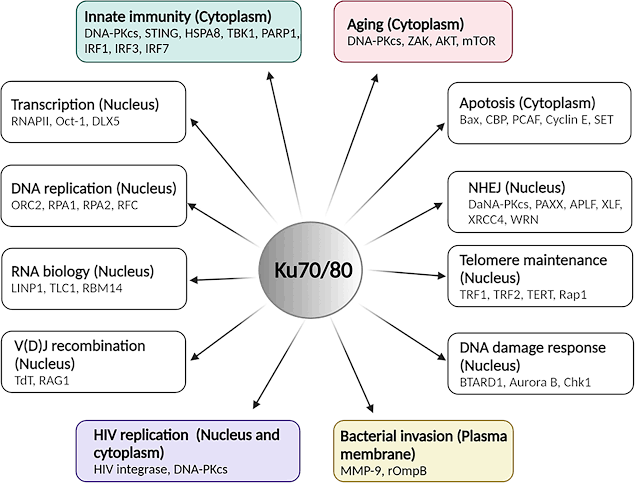 Ku70 ku80,迄今发现的作用(参与艾滋病病毒的复制和向患者基因组整合)
Ku70 ku80,迄今发现的作用(参与艾滋病病毒的复制和向患者基因组整合)
 Ku70 发生作用的蛋白
Ku70 发生作用的蛋白
无论是ku70 与NOX2, 还是Ku80与COX2 都与肿瘤生长有关。我们的工作表明,Ku70抑制NOX2是避免肿瘤细胞出现过多的氧游离基,从而帮助肿瘤长生不死!同时,Ku70自身还与细胞凋亡分子BAX结合,阻止它启动线粒体凋亡途径,使肿瘤细胞活下去!而Ku80促进COX2基因转录则顺势“炎性”反应,帮助肿瘤成长。
可见Ku70和ku80 除防止DNA断裂后被“大幅”降解之外并没干多少好事,包括降低氧游离基水平保护肿瘤细胞不发生副凋亡和坏死;通过“束缚”BAX避免凋亡性死亡;和炎性因子一道帮助肿瘤生长。即使是在可以帮助断裂的DNA免被大幅降解的过程,它还可以帮助通过“非同源末端连接”(non-Homologous End Joining, NHEJ)造成染色质重排,移位等等。
当然Ku蛋白的“本家兄弟”很多也表现出了“正能量”,会参与端粒维护,制造抗体分子的V(D)J 重排以及减数分裂过程中重组,包括“交配型”(Mate type 的Mating SWitch)。这些都对基因组稳定和基因多型性方面有贡献。
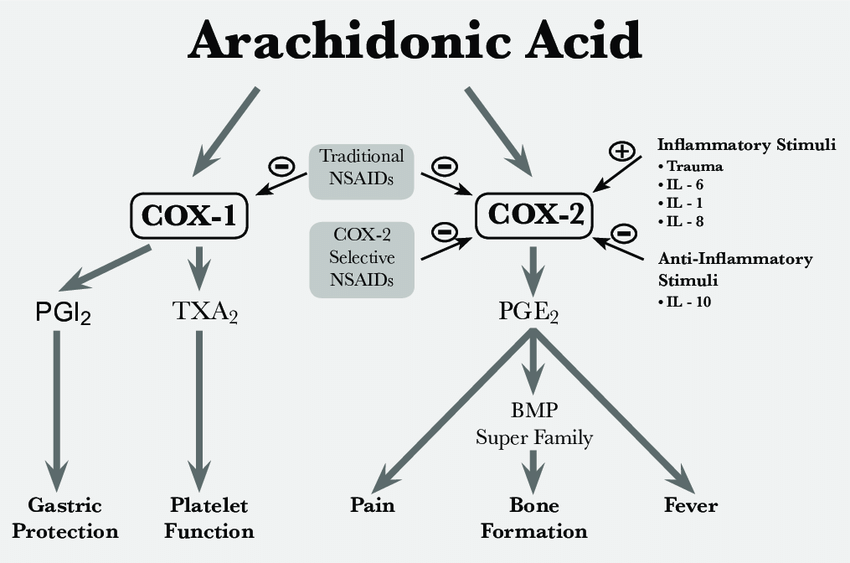 炎性过程中的COX
炎性过程中的COX
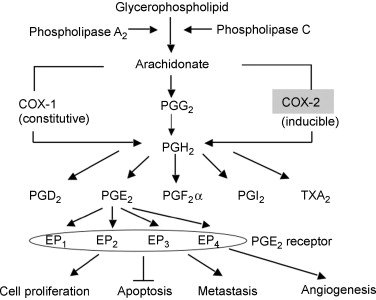 COX制造的前列腺素与肿瘤
COX制造的前列腺素与肿瘤
14、神秘的缠绕-不能捆绑不能断裂
DNA分子的生物活性取决于其拓扑结构,由碱基对沿着“中心轴”螺旋“盘绕”,这是拓扑连接子数的Twist, 大约10个碱基对一个Twist。如果DNA呈rcDNA或干脆呈linear DNA则不具进一步“缠绕”能力,即Writhe为0。这样的分子因不能被细胞内的拓扑异构酶加工,更重要的是体内的类似外切酶V的DNA外切酶会把线型DNA彻底降解,与此同时,碱基错配对蛋白也会干扰外源DNA分子通过同源重组向所在细胞的基因组进行整合。因此DNA外切酶V和碱基错配对识别蛋白一道发挥着“分子屏障”的作用,最受影响的是“自然转化”和人工转化线型DNA。
所谓“自然转化”是自然界中死亡的生物细胞的DNA分子“进入”活着的生物细胞,并试图向接收它的基因组整合的过程。这既是转基因作物有可能导致基因污染的生物学基础,也是“所有生物共享DNA”说法的依据。
自然界中主要存在两种DNA形式,一种是环状,一种是线性。前者常见于原核细胞、质粒、DNA病毒和动物细胞内的线粒体DNA(mtDNA)和叶绿体DNA,以及癌症患者的“癌基因”;绝大多数的动植物的基因组的DNA分子为线性。
这些DNA分子通过“成环”或通过CTCF、Cohesin和Codensin工具把长线型DNA分子“分段成环(Looping)”,为Writhe(缠绕, 这些缠绕所储存的能量用于DNA代谢,右旋,负超螺旋所储存的能量比左手正超螺旋的小,除了某些嗜热故细菌外,绝大多数生物的DNA均为右手负缠绕)的引入提供了可能。
但在DNA复制过程中,复制叉内的前导链和后随链受复制叉前“正超螺旋”的影响会向复制叉后传递,使得两条子链“缠绕”。这妨碍其接下来向子细胞内进行的“核分裂”。为此Cohesin 蛋白会把两条新生DNA 链“捆绑”,以避免彼此缠绕在一起。
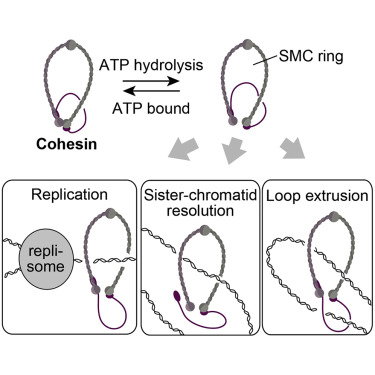 ATP ase Cohesin的
ATP ase Cohesin的
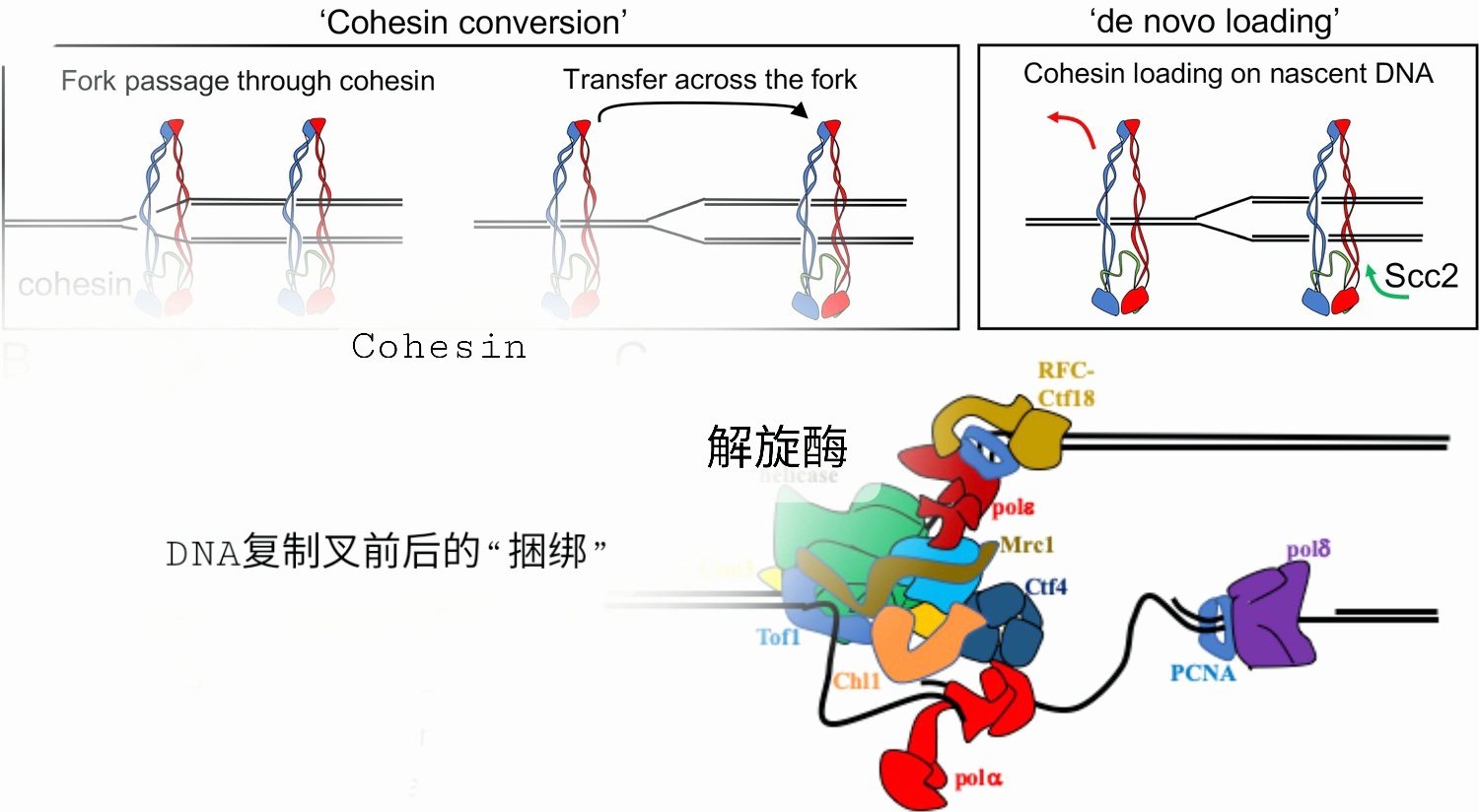 复制叉前后的捆绑
复制叉前后的捆绑
15、基因组的维修工-从SbcCD到MRN
从原核细胞到人类细胞均存在MR复合体,MR复合体由具有3'-5'外切酶,单链DNA内切酶活性的Mre11(Meiotic Recombination Exonuclease 11, Mre11)与具有ATPase活性的类SMC蛋白(Structural Maintenance of Chromosome, 或 Maintenance of small Chromsome(指啤酒酵母的第4号小染色体, SMC,类似人类基因组中的性染色体 Y)Rad50 (Rad50 编号以下的均为核酸酶)组成。其中,2个Rad50通过锌离子在“Zinc-hook” 部位形成“Y”型结构,而2个MRE11 分别与2个Rad50的N端和C端ABC结构域结合。Rad50 的2个ABC结构域分别具有ATPase结合和水解能力,分别称为“Walker A”和“Walker B”,而Mre11 大多数时候从DNA端部15~20个碱基处内切DNA, 之后利用其3'-5'外切酶降解,直到在另一条链上修剪出一段(~700bp)的单链供Rad51催化同源重组,或修剪出小段DNA 在DNA双链断裂处“互补”,之后被DNA连接酶IV连接(据说这个连接酶来源于拓扑异构酶,因为含有Toprim)。MR在原核大肠杆菌为SbcCD,与上述包括人在内的真核生物细胞的MR高度保守。
同源重组
 大肠杆菌依赖Chi的同源重组
大肠杆菌依赖Chi的同源重组
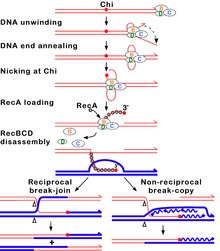
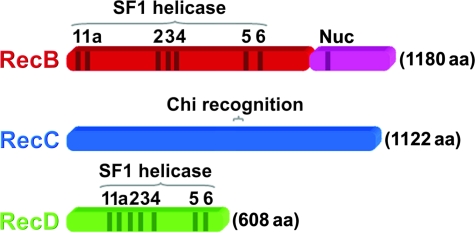
尽管大肠杆菌的SbcCD与真核细胞的MR结构保守,而真核生物细胞内的“同源重组”依赖MR产生“可重组”3'端,但在大肠杆菌细胞中,同源重组3'端的产生却并不是SbcCD, 而是使用一种依赖ATP的RecBCD, 又称“外切酶V”。RecBCD由RecB、RecC和RecD3个亚基组成,其中,RecB具有相对较弱的3'-5'解旋酶活性和外切酶活性,RecD则拥有5'-3'解旋酶活性,而RecC负责与一个被称为“chi”的“GCTGGTGG”识别,而chi可以激活RecB的核酸酶活性。
在催化过程中,RecB中的核酸酶可以根据反应条件以不同的方式起作用,特别是Mg2+离子和ATP浓度的比率。
(1) 如果ATP过量,RecB的内切酶将在Chi(具有3'端延伸的链)一侧内切,继续解旋打开双链,并产生一条3’“可重组”的尾巴,末端附近有Chi。该尾部可与RecA蛋白结合,有利于RecA催化的与其具有完整同源的DNA双链发生链侵入(strand invasion)当RecBCD到达DNA末端时,RecBCD所有三个亚单位都会分开,酶保持失去活性一小时或更长时间, 此时,与第一个Chi作用的RecBCD分子不会再攻击另一个DNA分子;


![]()
RecBCD三个亚基互动
 RecBCD的解旋酶活性
RecBCD的解旋酶活性
 结构域
结构域
(2) 如果Mg2+离子过量,则RecBCD内切两条DNA链,尽管5'尾的切割频率较低。当RecBCD在3'末端链上遇到Chi位点时,暂停,3'尾部消化减少。 当RecBCD恢复活性时,转而切割相反的链(即5′尾)。并将RecA蛋白加载到3′端链上。 在一个DNA分子上完成反应后,酶迅速攻击第二个DNA分子,在第二个DNA分子上发生与第一个DNA相同的反应。
尽管这两种反应都未通过细胞内DNA的分析得到证实,由于反应中间体的瞬时性质,遗传证据表明,第一种反应更接近于模拟细胞中的反应。 例如,Chi的活性受其3'侧核苷酸的影响,无论是在细胞中还是在与过量ATP的反应中,但与过量Mg2+的反应中,缺乏可检测核酸外切酶活性的RecBCD突变体在细胞中保持高Chi热点活性,并在细胞外的Chi处形成缺口。
细胞中一个DNA分子上的Chi位点降低或消除另一个DNA上的Chi活性,可能反映了在过量ATP和Chi处DNA缺口条件下体外观察到的RecBCD的Chi依赖性分解。
在两种反应条件下,3’链在Chi下游保持完整。然后通过RecBCD将RecA蛋白主动加载到3’尾部。 在某些未确定的点,RecBCD与DNA解离,尽管RecBCD可以解开至少60kb的DNA而不脱落。
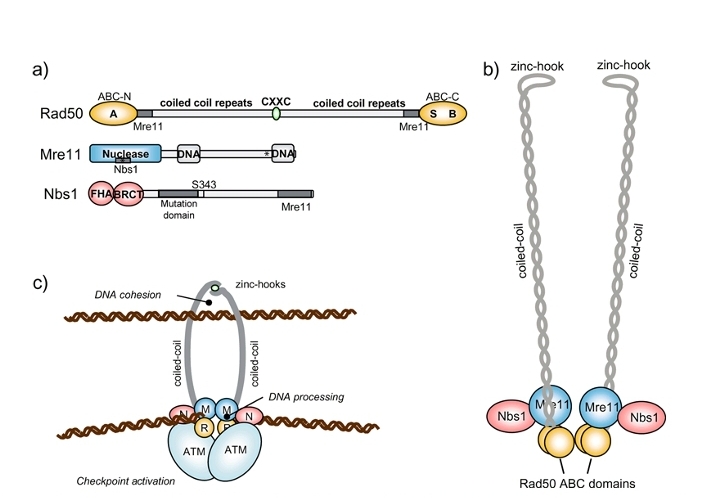 真核生物的MRN
真核生物的MRN
大肠杆菌中的SbcCD结构上与真核生物的MR相似
很显然,在同源重组过程中,大肠杆菌利用RecBCD依赖Chi序列(GCTGGTGG)调节其5'-3'和3'-5'外切酶活性,修剪出约70bp的可重组末端,而真核生物细胞则利用MRN做这个事,因为真核细胞内没有类似RecBCD的机制。
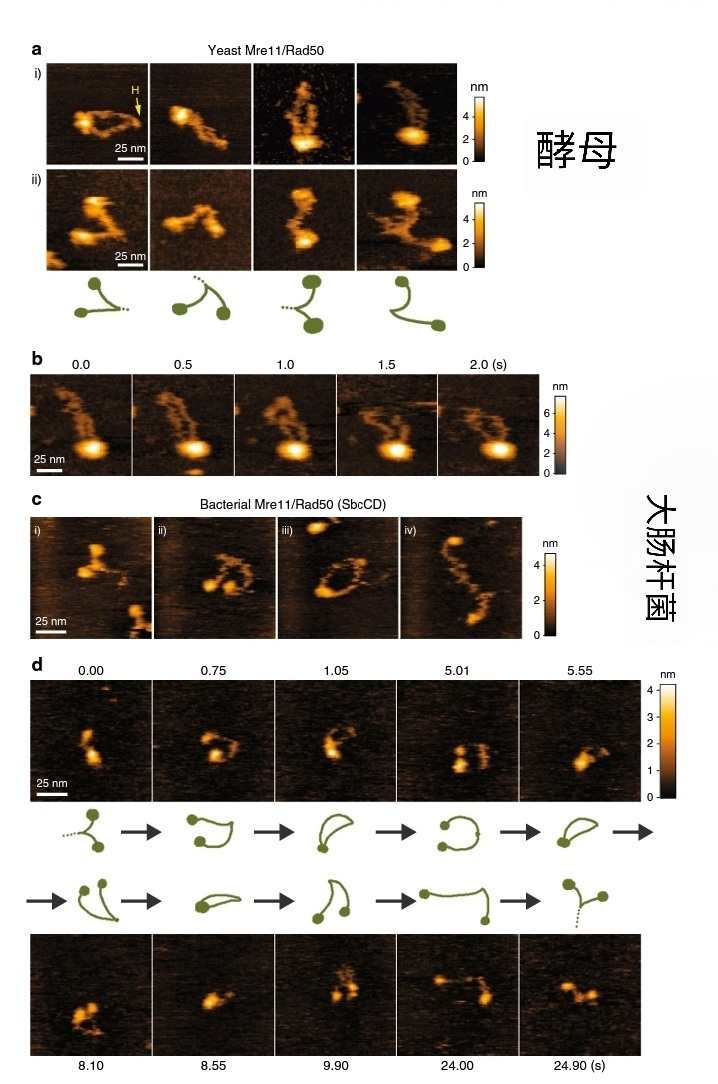
大肠杆菌和酵母的MR
 人类的MRN
人类的MRN
事实上,在组织结构上,大肠杆菌的SbcCD与真核细胞的Mre11-Rad50一致。但大肠杆菌的SbcCD更多用于同源重组和DNA复制的后随链和前导链切断含有非B型二级DNA结构的DNA外,似乎并不参与重组酶RecA 与单链DNA(ssDNA)组装成RecA-ssDNA, 而MRE11-Rad50确实不止表现出类似SbcCD的作用,同时它与Nbs1(人类Nijmegen 综合征, 全称 Nijmegen Breakage Syndrome )/Srs2(酵母)以MRN三聚体的形式参与同源重组过程的“可重组末端”的加工,在这个过程中可能也需要ExoI(外切酶I)助力。
因此,真核细胞内的MRN同时“扮演”原核细胞内的RecBCD(外切酶V)和SbcCD的“角色”。
16、解旋酶缺陷与人类疾病
据分析,迄今已经在人类细胞内发现了至少95种非冗余解旋酶,包括64种RNA解旋酶(helic“螺旋”+ase“酶”,翻译为“解旋酶” 是基于大多数此类酶所发挥的催化效果对应的翻译), 和31种DNA解旋酶。
一、DNA解旋酶超家族
其中的31种DNA解旋酶大致可分为6个超家族(Superfamily, SF)。
分别为:
1、 SF1
SF1(分 IA, 催化方向3'-5'和IB, 催化方向 5'-3')。这一组中,解旋酶既可以具有3'-5'(SF1A亚家族)催化活性,也有些具有5'-3'(SF1B亚家族),这里所述及的“方向”是指解旋酶沿着一条链“移动”的方向(催化过程中移动的极性/方向性)。代表解旋酶包括:革兰氏阴性细菌中的Rep解旋酶(负责清除与DNA结合的蛋白、UvrD(参与碱基错配对和核苷切除修复过程中的“开链”)以及PcrA解旋酶。SF1B组中以参与同源重组的RecD和参与修复的Dda解旋酶为代表。这些解旋酶在结构上均含有一个类似RecA折叠(RecA fold)的核心。
2、 SF2
这个超家族的解旋酶参与多种细胞过程。它们在结构上均含有9个保守基序,分别为Q、I、Ia、Ib和II~VI。这组解旋酶大多是含有“DEAD”盒(box)的RNA解旋酶。此外,RecQ样(RecQ-like)家族和Snf2样酶均被分在此超家族。SF2中的成员大多数为A型(3'-5'移位),少数如XPD家族例外。SF2的成员结构上也含有类似RecA的折叠核心。
3、SF3
SF3由主要由小型DNA病毒和一些大型核质DNA病毒编码的AAA+解旋酶(ATP associated multiple Activity, AAA+)组成。
它们具有3'-5'移位方向性,这意味着它们都是A型解旋酶。了解最清楚的SF3解旋酶是乳头状瘤病毒E1解旋酶。
4 、SF4
所有SF4成员都属于B型解旋酶(5’-3’)。在结构上含有一个RecA折叠。其中,研究最多的是来自噬菌体T7的gp4。
5、SF5
转录终止蛋白Rho属于SF5。这个家族的成员结构上也有一个RecA折叠。
6、SF6
SF6超家族成员含核心“AAA+”,但不算在SF3分类中。了解最多的SF6成员包括:小染色体稳定维护解旋酶MCM,以及大肠杆菌细胞中负责拆分“Holliday juncture ”的RuvA、RuvB和RuvC。
从结构上含有RecA fold 、P环(P-loop, 传递ATP水解后产生的构象改变,有利于移动)或“Walker”基序(具有ATPase活性)。这些是解旋酶能够“移位” 的结构基础。
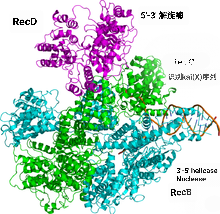 RecD解旋酶和RecB解旋酶组成
RecD解旋酶和RecB解旋酶组成
二、DNA解旋酶缺陷与人类疾病
DNA解旋酶基因突变/异常大多情况下通过影响基因组稳定性和基因表达出现症状。以下数例为代表。
1、ATR-X 综合征(syndrome)
ATRX( X-linked-alpha-thalassaemia-mental retardation,X-连锁α-地中海贫血-智力低下)基因位于X染色体Xq13.1-q21.1,编码一个属于SNF2超家族的ATP依赖性解旋酶ATRX(也称为XH2、XNP),该解旋酶负责染色质重塑(Chromatin Remodeling)、基因调控和rDNA(核糖体DNA甲基化等。这些功能有助于防止细胞凋亡,对大脑皮层的大小具有调节作用,同时,有助于海马和皮层神经元细胞的存活,因此影响记忆和学习。有意思的是该解旋酶基因位于着丝粒周围的异染色质中,且与异染色质蛋白(HP1)1结合。
患者主要表现包括:小头畸形、骨骼和面部异常、智力低下、生殖器异常、癫痫、语言使用受限及α-地中海贫血等。常见突变大多数位于锌指结构域和解旋酶结构域,包括无义点突变、缺失、移码突变等等。突变基因或许影响了其下游的血红蛋白基因的表达。
2、布鲁姆综合征(BS)、罗斯蒙德-汤姆森综合征(RTS)和沃纳综合征(WS)
布鲁姆综合征(BS)、罗斯蒙德-汤姆森综合征(RTS)和沃纳综合征(WS)的突变基因属于 RecQ家族,分别为BLM、RECQL4和WRN。这些解旋酶的缺陷和/或突变显示异常的基因重组和/或DNA复制,这导致染色体不稳定性和增殖能力的总体降低,因此受累基因BLM、RECQL4和WRN中的突变在调节同源重组中发挥作用。
布鲁姆综合征
布鲁姆综合征的典型病理特征为癌症易感,达到发病早,平均发病年龄为24岁左右。BLM细胞表现出姐妹染色单体(SCE)之间的高频率互换和过度染色体损伤。有证据表明,BLM在修复复制分叉处中断的DNA复制中发挥作用;
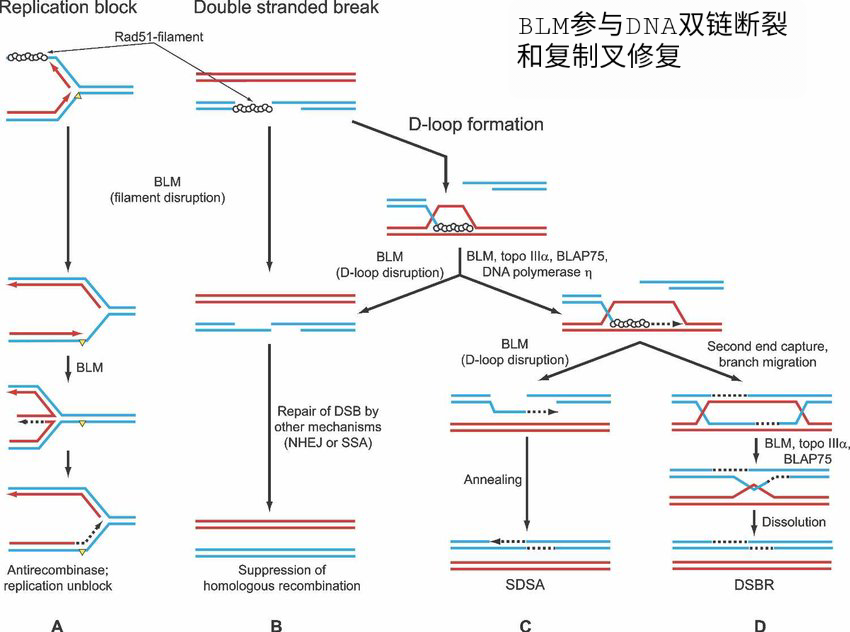 BLM解旋酶
BLM解旋酶
Werner综合征
这种疾病的典型病理特征为早衰,包括动脉粥样硬化和骨质疏松症以及其他年龄相关疾病的早期发病、肉瘤的高发,以及在生命的第4至第6个十年中经常因心肌梗死或癌症而死亡;
WRN的细胞表现出染色体断裂和易位。细胞的生殖寿命缩短,以及染色体组分大量缺失,导致基因组不稳定;
Rothmund-Thomson综合征
该病也被称为先天性骨肉瘤,其特征是过早衰老、皮肤和骨骼异常、皮疹、骨肉瘤、青少年白内障和易患癌症(如骨肉瘤)。在Rothmund-Thomson综合征患者的细胞中发现了导致基因组不稳定性的染色体重排。
3、Cockayne综合征(CS)和毛发硫营养不良症(TTD)
这两类综合征与解旋酶XPD, 该解旋酶属于SF2超家族,拥有5'-3'DNA解旋酶活性。
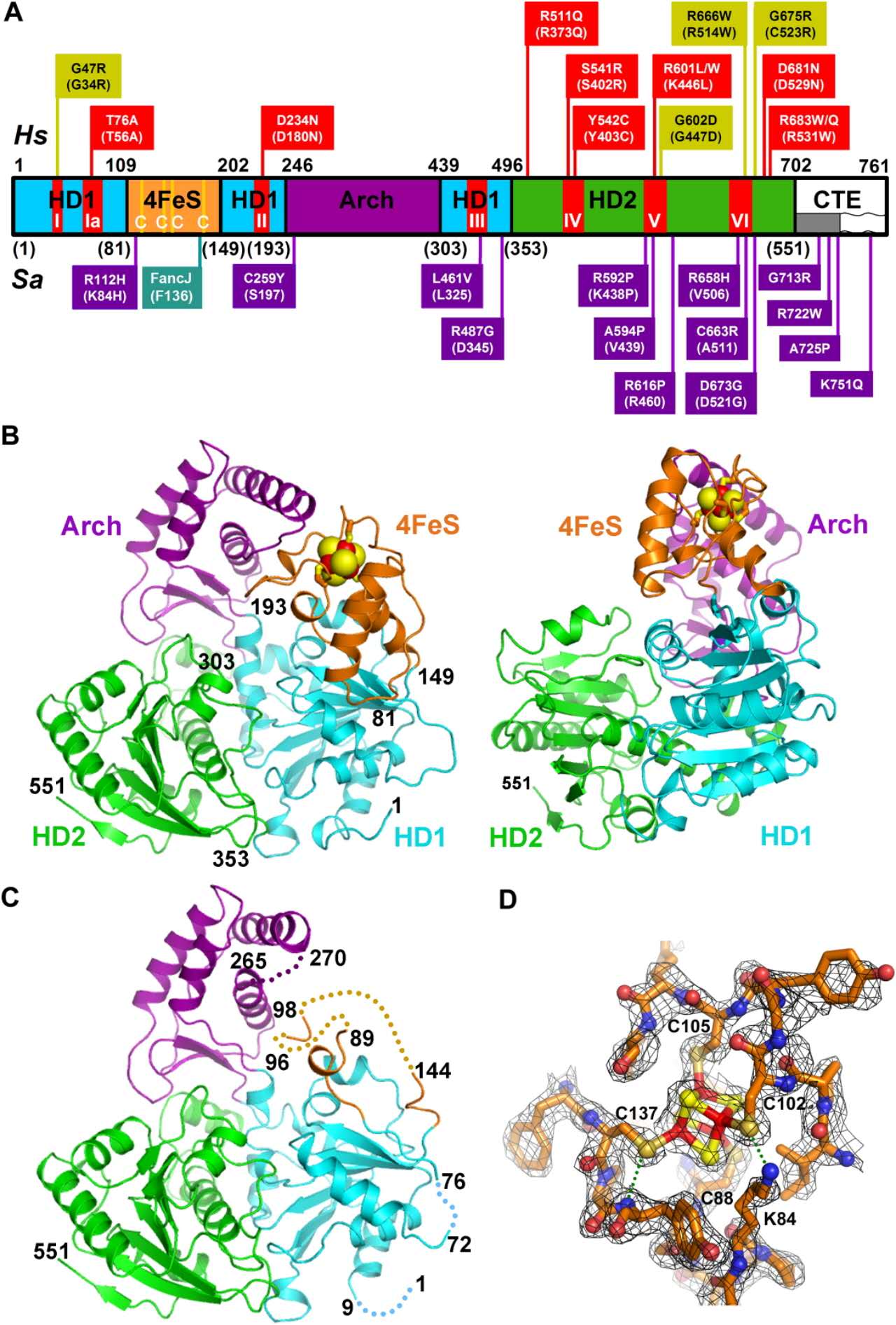 XPD解旋酶的结构及突变
XPD解旋酶的结构及突变
https://m.sciencenet.cn/blog-218980-1354367.html
上一篇:科学遐想-神秘的缠绕(7--12)
下一篇:iPSCs 和CRISPR-Cas9何时能梦圆医疗实践?