博文
“郁金香树”~鹅掌楸属植物
||
鹅掌楸属(Liriodendron)植物是木兰类(magnoliids)木兰科(Magnoliaceae)鹅掌楸亚科(Liriodendroidae)的单属物种。鹅掌楸属包含一对姊妹种:一种自然分布于东亚地区,即鹅掌楸(L. chinense (Hemsl.) Sargent.)[1];一种自然分布于北美东部地区,即北美鹅掌楸(L. tulipifera Linn.)[2]。
鹅掌楸属植物是第三纪孑遗植物,该属植物曾有10余种,化石资料显示其在第三纪时期曾广泛分布于北半球高纬度地球。而后,鹅掌楸属经历过晚第三纪时期的降温及第四纪冰川后,现仅存两种,在各大陆上的中、低纬度地区分布,且自然分布与第三纪两个孑遗植物区系的分布范围一致[8]。尽管鹅掌楸属植物也曾在欧洲的高纬度地区有所分布,然而欧洲东西走向的山脉在第三纪晚期及第四纪冰川时期阻断了鹅掌楸属植物南迁的道路,最终导致了该类植物在欧洲的灭绝[3, 9]。
1. 鹅掌楸与北美鹅掌楸
鹅掌楸属植物花大而艳丽,形似郁金香,所以又名郁金香树(tulip tree),而鹅掌楸属名Liriodendron实则源自于希腊语中的lerion,lily和dendron,意指开着像百合那样花朵的树木[7]。鹅掌楸属植物现存仅两种,鹅掌楸与北美鹅掌楸,均属于高大乔木,二者大概在中新世中期开始分化,至今已经分化了约1,000万年到1,600万年[3]。鹅掌楸与北美鹅掌楸之间的形态学特征大体相似,但从叶形、树皮及花等表型性状可明显区分这两个种[11]。鹅掌楸叶一般3裂,树皮裂缝不明显,花被片绿色,有黄色纵条纹,基部蜜腺不发达;而北美鹅掌楸叶一般5裂,树皮纵裂纹较深而明显,花被片绿黄色,基部内侧有丰富的橙黄色蜜腺。

图1 鹅掌楸(左)与北美鹅掌楸(右)的花

图2 鹅掌楸盛花期花的解剖(比例尺:2 cm;排布参考微博“璐角海棠”)

图3 北美鹅掌楸盛花期花的解剖(比例尺:2 cm)
北美鹅掌楸广泛分布于美国东部至加拿大东南部,且在当地林分中常占优势地位;相比而言,鹅掌楸在中国的分布呈星散间断分布,且天然更新不良,已被列入我国国家二级珍稀濒危保护植物[1]。相比于鹅掌楸,北美鹅掌楸不仅在北美地区依靠种子的天然更新能力强,自然分布广而密集,且在美国的栽培历史较早,为优良的工业原料林树种。早在1663年,北美鹅掌楸就已经开始被用于人工栽培[12]。随后,北美鹅掌楸种子园的建立极大满足了当地和区域性栽培苗的供应和需求[13, 14]。其中,1966年在田纳西Knoxville建立的种子园,共包含31个基因型(或无性系),每个基因型繁殖100嫁接苗;1976年在南卡罗来纳Clemson建立的种子园,是由克莱姆森大学的Roland E. Schoenike博士从17500英亩的克莱姆森试验林中选择的150优树进行嫁接形成的。现在,Clemson种子园中还剩165株鹅掌楸,其中大部分是北美鹅掌楸,还有少数几株鹅掌楸[15]。北美鹅掌楸的木材应用广泛,可供建筑、家具、造纸用材等[16]。同时,北美鹅掌楸的花蜜十分丰富,被认为是蜜蜂的重要蜜源植物,同时也是一些野生动物的食物来源;此外,鹅掌楸还是城市绿化的重要遮阴树[17, 18]。另外,在北美鹅掌楸木材和叶片上提取的代谢化合物具有非常重要的医用价值,如倍半萜具有抗肿瘤活性,且能抗食草动物的啃食[19];而生物碱类的代谢物则具有一定程度上的抗菌作用[20]。
2. 鹅掌楸杂交育种
我国已故著名植物育种学家叶培忠教授自上个世纪60年代利用20世纪30年代引种至南京明孝陵森林公园的一株北美鹅掌楸的花粉与鹅掌楸进行种间杂交试验,并于1973年正式发表文章报道了亚美鹅掌楸杂交育种的成功[4]。

图4 鹅掌楸属植物种间杂交制种

图5 杂交鹅掌楸的花

图6 杂交鹅掌楸盛花期花的解剖(比例尺:2 cm)

图7 杂交鹅掌楸聚合翅果(比例尺:2 cm)
育成的杂交鹅掌楸(学名:亚美马褂木;拉丁名:Liriodendron sino-americanum P. C. Yieh ex Shang et Z. R. Wang)存在较强的杂交优势,生长迅速、树形挺拔、干形通直、叶形奇特、花色艳丽,是重要的造林树种和道路景观树种[5, 6]。

图8 杂交鹅掌楸的幼苗、成年大树及叶片表型
美国北卡罗来纳大学的Clifford R. Parks等人于1973年对鹅掌楸与北美鹅掌楸进行了种间和种内杂交试验,并于7年后测定了杂种后代的生长量,结果显示种间杂交比种内杂交明显生长占优[2]。同样,南京林业大学育种组于1978年在南京林业大学树木园建立了鹅掌楸杂种试验林,其20年生的生长量数据(表1)也显示了杂交鹅掌楸具有十分明显的杂种优势[21]。
表1 鹅掌楸属种间杂交的生长表现
性状 | 杂交鹅掌楸 | 鹅掌楸 | 北美鹅掌楸 | 杂种优势(%) |
树高(m) | 19.17 | 15.52 | 16.22 | 20.79 |
胸径(cm) | 25.71 | 13.56 | 16.77 | 69.59 |
材积(m3) | 0.460 | 0.107 | 0.170 | 232.13 |
注:引自王章荣等,鹅掌楸属树种杂交育种与利用(第二版),P128,2016,中国林业出版社。
尽管杂交鹅掌楸有着诸多优越性,然而天然条件下鹅掌楸自然结实率极低,人工杂交费时费力且受季节限制,严重阻滞了杂交鹅掌楸的推广应用[22]。尽管2002年,南京林业大学陈金慧等以杂交鹅掌楸春季新芽为材料,进行不定芽的诱导和植再生,成功构建了一套杂交鹅掌楸组培快繁的流程,但杂交鹅掌楸仍存在不定芽生根困难的问题[23]。1986年,美国乔治亚大学的Merkle成功利用北美鹅掌楸的未成熟种子的胚和胚乳构建了体胚发生体系[24]。之后,1990年,Merkle利用3个木兰属(Magnolia)物种未成熟种子的胚和胚乳成功构建了体胚发生体系[25]。接着,在1993年Merkle利用杂交鹅掌楸未成熟的种子,成功构建了体胚发生体系[26]。
我国南京林业大学林木遗传与生物技术教育部重点实验室,林木细胞工程团队(负责人:施季森教授、陈金慧教授)于2003年成功以杂交鹅掌楸幼胚为外植体,建立了杂交鹅掌楸的体细胞胚胎发生技术,并由此成功开发了杂交鹅掌楸的快速成苗体系,首次实现了工业化的规模生产,开辟了杂交鹅掌楸产业化开发的新途径[27]。接着,该团队又进一步对杂交鹅掌楸体胚系统的遗传稳定性进行了研究,显示杂交鹅掌楸胚性细胞系的体胚发生具有长期保存能力,为杂交鹅掌楸体胚植的商业化应用提供了理论基础[28]。之后数年间,该团队对杂交鹅掌楸体胚发生体系进行了一系列的优化,并对获得的体胚苗进行大田试验、测定和选育,申请获得了杂交鹅掌楸林木良种并进行了推广造林,极大的推进了杂交鹅掌楸在我国的推广应用[29-33]。

图9 杂交鹅掌楸体细胞胚胎发生体系
除了在鹅掌楸属植物的细胞工程育种领域取得了巨大的成就外,该团队于2019年首次破译了鹅掌楸全基因组[34],并基于此基因组对鹅掌楸的生殖繁育性状、逆境胁迫、倍性育种等进行了系统研究[35-40]。同时,该团队还基于成熟的杂交鹅掌楸体胚发生体系,开发了高效的遗传转化体系[41];目前,正在构建基因编辑体系并成功获得了基因编辑的再生植株(未发表)。相信在不久的未来,鹅掌楸属植物在全基因组选择育种、转基因育种、基因编辑育种等新兴分子精准育种领域将大有可为。

图10 杂交鹅掌楸稳定遗传转化体系

图11 鹅掌楸野生型与多季节开花突变体slb1的开花习性对比

图12 鹅掌楸野生型需8-10年开花 vs slb1自交子代4个月开花

图13 二倍体 vs 四倍体杂交鹅掌楸
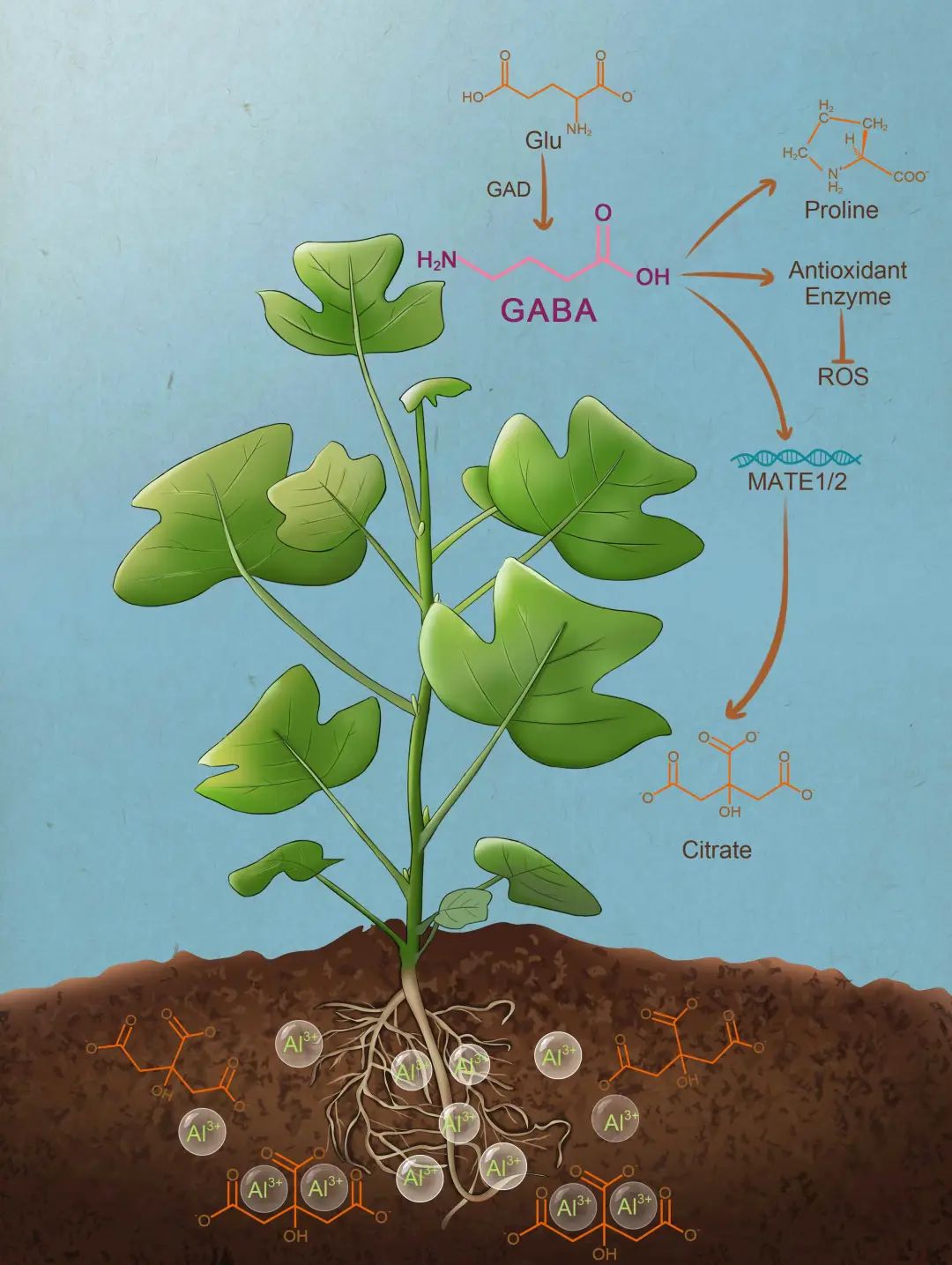
图14 GABA介导的杂交鹅掌楸铝离子胁迫抗性
参考文献
[1] 郝日明和贺善安. 鹅掌楸在中国的自然分布及其特点. 植物资源与环境, 1995, 4(1): 1-6.
[2] Parks C. R., Miller N. G., Wendel J. F., et al. Genetic-divergence within the genus Liriodendron (Magnoliaceae). Ann Mo Bot Gard, 1983, 70(4): 658-666.
[3] Parks C. R. and Wendel J. F. Molecular divergence between Asian and north-American species of Liriodendron(Magnoliaceae) with implications for interpretation of fossil floras. Am J Bot, 1990, 77(10): 1243-1256.
[4] 南京林产工业学院林学系育种组. 亚美杂交马褂木的育成. 林业科技通讯, 1973, 12: 10-11.
[5] 李火根和施季森. 杂交鹅掌楸良种选育与种苗繁育. 林业工程学报, 2009, 23(3): 1-5.
[6] 向其柏和王章荣. 杂交马褂木的新名称—亚美马褂木. 南京林业大学学报(自然科学版), 2012, 36(2): 1-2.
[7] Coombes A. J. The A to Z of Plant Names: A Quick Reference Guide to 4000 Garden Plants, 2012, Timber Press, Portland.
[8] Milne R. I. Northern Hemisphere plant disjunctions: a window on tertiary land bridges and climate change? Annals of botany, 2006, 98(3): 465-472.
[9] Tiffney B. H. The Eocene North Atlantic land bridge: its importance in Tertiary and modern phytogeography of the Northern Hemisphere. Journal of the Arnold Arboretum, 1985, 66(2): 243-273.
[10] Nie Z. L., Wen J., Azuma H., et al. Phylogenetic and biogeographic complexity of Magnoliaceae in the Northern Hemisphere inferred from three nuclear data sets. Mol Phylogenet Evol, 2008, 48(3): 1027-1040.
[11] 刘玉新, 王章荣, 黄淑婧, 等. 杂交马褂木形态特征识别要点. 湖北林业科技, 2014, 43(3): 74.
[12] Bonner F. T. and Russell T. E. Liriodendron tulipifera L. Yellow-poplar, 1974, In: Seeds of woody plants of the United States. USDA For Serv, Washington, DC, 508–511.
[13] Franklin C., Brown J. and Weingartner D. Wind damage to a yellow-poplar seed orchard. Tree Planters’ Notes, 1976, 27(1): 3-4.
[14] Eyvind T. Tree Breeding at the University of Tennessee 1959-1975, 1976, Bulletins.
[15] Zhang X., Carlson A., Tian Z., et al. Genetic characterization of Liriodendron seed orchards with EST-SSR markers. Journal of Plant Science and Molecular Breeding, 2015, 4(1): 680-692.
[16] Williams R. S. and Feist W. C. Durability of yellow-poplar and sweetgum and service life of finishes after long-term exposure. For Prod J, 2004, 54: 96-101.
[17] Zhou Y., Li M., Zhao F., et al. Floral nectary morphology and proteomic analysis of nectar of Liriodendron tulipiferaLinn. Front Plant Sci, 2016, 7: 826.
[18] Beck D. E. Liriodendron tulipifera L. Yellow-poplar, In: Silvics of North America: 2, Harwoods. U.S. Department of Agriculture, Forest Service, Washington, DC, 406-416.
[19] Moon M. K., Oh H. M., Kwon B. M., et al. Farnesyl protein transferase and tumor cell growth inhibitory activities of lipiferolide isolated from Liriodendron tulipifera. Arch Pharm Res, 2007, 30(3): 299-302.
[20] Hufford C. D., Funderburk M. J., Morgan J. M., et al. Two Antimicrobial Alkaloids from Heartwood of Liriodendron tulipifera L. J Pharm Sci, 1975, 64(5): 789-792.
[21] 王章荣. 中国马褂木遗传资源的保存与杂交育种前景. 林业实用技术, 1997, 9: 8-10.
[22] 季孔庶. 杂交鹅掌楸的无性繁殖. 南京林业大学学报(自然科学版), 2005, 29(1): 83-87.
[23] 陈金慧, 施季森和诸葛强. 杂交鹅掌楸的不定芽诱导及植株再生. 植物生理学报, 2002, 38(5): 459.
[24] Merkle S. A. and Sommer H. E. Somatic embryogenesis in tissue cultures of Liriodendron tulipifera. Can J For Res, 1986, 16(2): 420-422.
[25] Merkle S. A. and Wiecko A. T. Somatic embryogenesis in three magnolia species. J Am Soc Hortic Sci, 1990, 115(5): 858-860.
[26] Merkle S. A., Hoey M. T., Watson-Pauley B. A., et al. Propagation of Lirlodendron hybrids via somatic embryogenesis. Plant Cell Tissue Organ Cult, 1993, 34(2): 191-198.
[27] 陈金慧, 施季森, 诸葛强, 等. 杂交鹅掌楸体细胞胚胎发生研究. 林业科学, 2003, 39(4): 49-53.
[28] 陈金慧, 施季森, 赵治芬, 等. 杂交鹅掌楸体胚系统的遗传稳定性研究. 南京林业大学学报(自然科学版), 2006, 30(6): 99-101.
[29] 陈金慧, 张艳娟, 李婷婷, 等. 杂交鹅掌楸体胚发生过程的起源及发育过程. 南京林业大学学报(自然科学版), 2012, 36(1): 16-20.
[30] 李雪萍, 边黎明, 陈金慧, 等. 整地方式对杂交鹅掌楸体胚苗及其亲本幼林生长的影响. 林业科技开发, 2012, 26(6): 59-61.
[31] 李雪萍, 赵胜杰, 边黎明, 等. 杂交鹅掌楸体胚苗及亲本种造林比较试验. 江苏林业科技, 2013, 39(6): 5-7.
[32] 陈金慧, 张艳娟, 吴亚云, 等. 植物磺肽素在杂交鹅掌楸体胚发生中的作用. 林业科学, 2013, 49(2): 33-38.
[33] 鲁路, 陆叶, 盛宇, 等. 不同活性炭对杂交鹅掌楸体胚发生的影响. 南京林业大学学报(自然科学版), 2016, 40(2): 59-64.
[34] Chen J., Hao Z., Guang X., et al. Liriodendron genome sheds light on angiosperm phylogeny and species-pair differentiation. Nat Plants, 2019, 5(1): 18-25.
[35] Sheng Y., Hao Z., Peng Y., et al. Morphological, phenological, and transcriptional analyses provide insight into the diverse flowering traits of a mutant of the relic woody plant Liriodendron chinense. Hortic Res, 2021, 8: 174
[36] Hao Z., Liu S., Hu L., et al. Transcriptome analysis and metabolic profiling reveal the key role of carotenoids in the petal coloration of Liriodendron tulipifera. Horticu Res. 2020, 7: 70.
[37] Wang P., Dong Y., Zhu L., et al. The role of γ-aminobutyric acid in aluminum stress tolerance in a woody plant, Liriodendron chinense × tulipifera. Horticu Res. 2021, 8(1): 80.
[38] Wu W., Zhu S., Xu L., et al. Genome-wide identification of the Liriodendron chinense WRKY gene family and its diverse roles in response to multiple abiotic stress. BMC Plant Biol. 2022, 22: 25.
[39] Wu W., Zhu S., Zhu L., et al. Characterization of the Liriodendron chinense MYB gene family and its role in abiotic stress response. Front Plant Sci. 2021, 12: 641280.
[40] Chen T., Sheng Y., Hao Z., et al. Transcriptome and proteome analysis suggest enhanced photosynthesis in tetraploid Liriodendron sino-americanum. Tree Physiol. 2021, tpab039.
[41] Li M., Wang D., Long X., et al. Agrobacterium-mediated genetic transformation of embryogenic callus in a Liriodendron hybrid (L. chinense × L. tulipifera). Front Plant Sci, 2022, 13: 802128.
(p.s. 文中图片版权归原作者所有,禁止一切商业目的用途;如需用于交流、学习、教学等目的,欢迎转载,但需注明来源,即“南京林业大学林木细胞工程课题组”)
https://m.sciencenet.cn/blog-3158122-1338701.html
上一篇:Molecular Plant:稻田四倍体杂草千金子基因组
下一篇:The Plant Cell:ABA合成基因上逐步式的染色质沉默是种子萌发的关键
全部作者的其他最新博文
- • Plant Physiology:CsMADS3促进柑果中的叶绿素降解和类胡萝卜素合成(华中农业大学)
- • Molecular Plant:LBD11-ROS反馈调节作用于拟南芥的维管形成层增殖和次生生长(浦项科技大学)
- • Science Advances:根结线虫通过调控植物的CLE3-CLV1模块,促进侵染进程(日本熊本大学)
- • The Plant Cell:拟南芥P小体组分通过对FLC的转录调控,影响开花时间(安徽农业大学)
- • Nature Communications:油菜素内酯参与植物营养生长期转变的分子机制解析(浙江农林大学)
- • Current Biology:光合作用产生的蔗糖驱动侧根“生物钟”(德国弗莱堡大学)