博文
代谢学人--Nature:凋亡的脂肪细胞也要发挥余温!
||
代谢学人
Nature:凋亡的脂肪细胞也要发挥余温!
撰文 | 郭文秀 生茂正 于剑
编辑 | 孟美瑶
校对 | 于剑

背景介绍
人体内,白色脂肪组织(WAT)负责将能量以脂肪的形式储存起来,棕色脂肪组织(BAT)则通过产热消耗能量来维持机体的代谢健康。肥胖和衰老均导致BAT的细胞数量减少,产生功能性萎缩和适应性产热障碍,这种功能变化是以BAT为中心的肥胖治疗过程的主要障碍。有文献认为,组织中脂肪细胞数量的增加可以通过α或β-肾上腺素诱导的细胞凋亡抑制来实现。然而,目前对BAT的凋亡了解不多。
BAT细胞与其他细胞类似,也会发生细胞凋亡,即Apoptosis,为维持内环境稳定,由基因控制的细胞自主有序的死亡,涉及一系列基因的激活、表达以及调控。该过程主要由Caspase家族和抑癌基因p53等多种基因介导调控。在凋亡发生后,细胞体积缩小,细胞质密度增加,线粒体膜电位消失,细胞通透性改变,形成凋亡小体,被其他吞噬细胞吞噬。作为一种细胞主动死亡的过程,棕色脂肪细胞凋亡是一个持续的过程,即使是在正常环境中,BAT组织也会发生缓慢的细胞凋亡,而去神经等慢性失活手段更会导致棕色脂肪细胞的活性和丰度进一步降低。但是棕色脂肪细胞凋亡的具体分子机制尚不清楚。
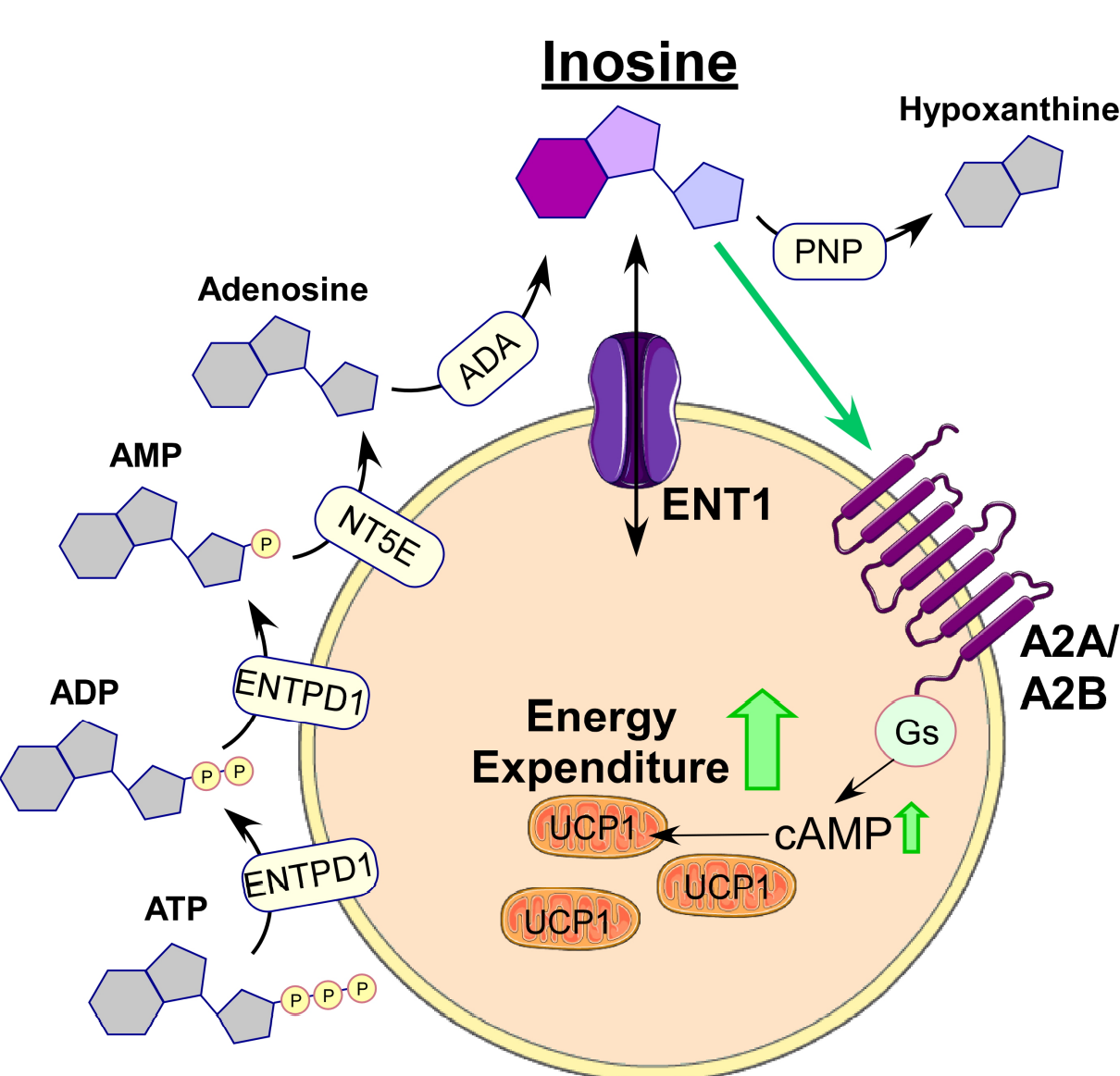
在先前的研究中,核苷的大量释放往往被认为是凋亡信号发生的标志性信号之一。核苷作为生物学上重要的分子,在核酸合成和能量代谢等方面发挥着巨大的作用,在细胞凋亡中的作用也不可小觑。有研究表明,嘌呤等核苷分子可以作为信号分子发挥相应功能。当细胞发生凋亡时,大量的ATP/UTP被释放,并作为“凋亡标志物”与巨噬细胞中的P2Y受体结合,从而募集到包括单核细胞、巨噬细胞和树突状细胞等的大量吞噬细胞,清除目标靶细胞完成凋亡反应。那么,BAT的凋亡是否也是通过核苷作为“寻找”信号,并以此来维护正常细胞的高效率工作呢?近期发表在Nature上的一篇题目为“Apoptotic brown adipocytes enhance energy expenditure via extracellular inosine”便针对这一问题进行了解答,研究发现肌苷(inosine)是BAT细胞凋亡过程中释放的代谢物,具有“取代我”的信号功能,可以促进产热并抵抗肥胖。

敲黑板啦!
1.棕色脂肪细胞凋亡会分泌嘌呤肌苷,其通过cAMP/PKA通路增强能量消耗。
2.ENT1敲除引起细胞外肌苷上调,从而增强产热基因表达和抵抗肥胖。
3.使用双嘧达莫抑制ENT1可增强脂肪细胞棕色化,提升能量消耗。
4.人群中存在SLC29A1的Ile216Thr突变使ENT1失活,并与较低的BMI有关。
研究结果
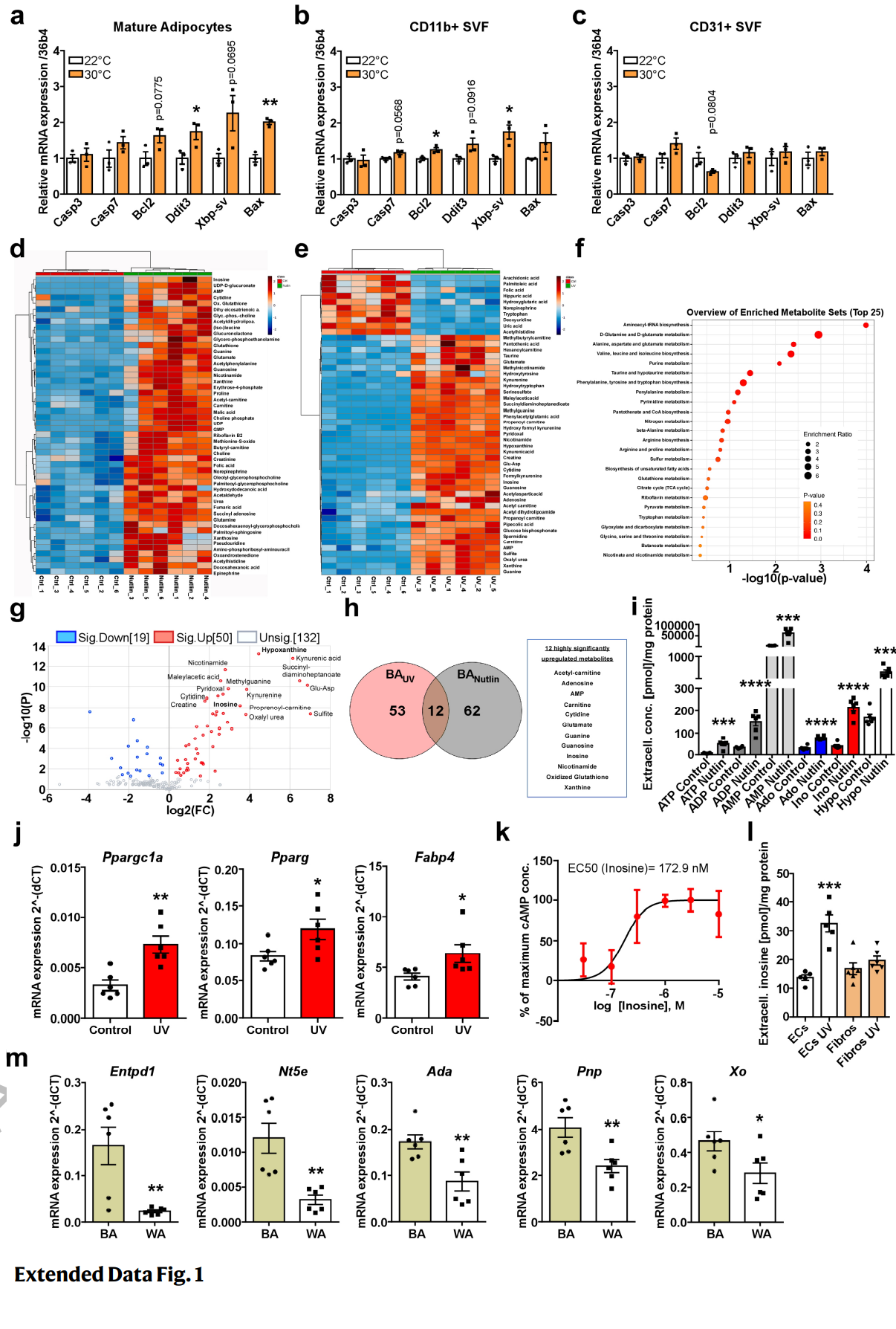
为了研究脂肪细胞凋亡在生理条件下是否会自然发生,作者将小鼠置于热中性(30°C)以使BAT失活。经过3天和7天的热中性处理后,发现BAT中凋亡的TUNEL阳性细胞显著增加(图1A)。为了确定发生凋亡的细胞类型,作者从30℃条件下饲养3天的小鼠BAT中分离出含量较高的细胞种类,包括成熟脂肪细胞、CD11b阳性免疫细胞和CD31阳性内皮细胞。与对照组(22°C)相比,热中性(30°C)组的成熟脂肪细胞中的凋亡标记物表达增加 (图S1A-C),包括促凋亡转录因子DNA损伤诱导转录因子3 (DNA damage-inducible transcript 3, dit3), X-box结合蛋白1 (X-box binding protein 1, Xbp1-sv)和BCL2相关X蛋白(BCL2 associated X protein, Bax)的剪接mRNA。值得注意的是,有研究表明即使在热中性暴露4周后,棕色脂肪细胞也会有类似的凋亡特征。这些结果表明,BAT的生理失活诱导适应性生理反应,促进产热脂肪细胞凋亡。
接下来,作者将小鼠前体脂肪细胞/间质血管组分(即SVF组分),分化为成熟的棕色脂肪细胞,并用紫外线(UV)照射以诱导Caspase(半胱天冬氨酸酶)依赖性凋亡,或用nutlin-3(小编注:MDM2的小分子抑制剂。细胞受刺激后,磷酸化的P53与MDM2/MDMX复合物解离,从而激活PUMA、BAX等凋亡基因的转录,引发细胞凋亡,从而抑制肿瘤发生。而nutlin-3的作用为抑制MDM2和p53的相互作用,并稳定p53蛋白防止其降解,从而进一步诱发凋亡)处理后以激活p53通路触发凋亡。这两种条件均可以显著激活脂肪细胞中的annexinV(一种标记凋亡信号的Marker)阳性信号,同时确保细胞膜的完整性(碘化丙啶阴性细胞)。在获取凋亡细胞后,作者使用非靶向代谢组学发现与棕色脂肪细胞凋亡相关的代谢产物,随后重点关注上调最高的代谢途径。通过检测细胞上清液中的代谢物的电荷质量、电荷比及其碎片模式并进行分析,发现nutlin-3诱导的细胞凋亡导致84种代谢物显著富集,13种代谢物显著减少(图1B)。在棕色脂肪细胞的上清液中总共检测到330种化合物,其中嘌呤能核苷酸(如琥珀酰腺苷、鸟苷、肌苷)在最显著上调的化合物中占100种(图S1D)。因此,代谢途径的定性富集分析显示,在nutlin-3诱导的凋亡过程中,嘌呤代谢是棕色脂肪细胞中变化最显著的代谢途径(图1C)。与此一致的是,紫外线照射诱导细胞凋亡也强烈影响BA的分泌组(图S1E),且嘌呤代谢途径在其中也起到了重要作用(小编注:排名为第五位。第一的为氨基酸-tRNA生物合成)(图S1F),分别有50种和19种代谢产物显著上调和下调 (图S1G)。在两种处理方式(nutlin-3或UV)中,有12种代谢产物显著上调,其中6种属于嘌呤能分子家族(图S1H)。
接下来通过对细胞外嘌呤能分子进行特异性分析,尤其是ATP衍生分子(ATP此前已被证明是其他细胞类型中主要的与凋亡相关代谢物)。在UV诱导的凋亡过程中,棕色脂肪细胞释放出一种特定的嘌呤分子模式,ATP水平显著升高(图1D),此外,另外三种显著增加的嘌呤分子,AMP、肌苷和次黄嘌呤,均达到了细胞外最高浓度(图1D)。在nutlin-3处理后,也观察到类似的模式(图S1I)。有趣的是,将健康的棕色脂肪细胞与凋亡的棕色脂肪细胞的上清孵育,可以显著增加健康脂肪细胞中产热标记物Ucp1和Ppargc1a以及成脂分化标记物 Pparg和Fabp4的基因表达 (图1e, 图S1J),这表明凋亡的棕色脂肪细胞发出替换信号以维持组织功能。

图1 | 肌苷对BAT代谢的促进作用
2、肌苷增强产热程序
鉴于凋亡棕色脂肪细胞上清液的刺激作用,作者分析了增强棕色脂肪细胞分化和产热程序的关键第二信使cAMP的浓度。为了确定介导这一效应的个体代谢物,作者重点研究了3种分泌最多的细胞外嘌呤(图1D)。腺嘌呤(AMP)和次黄嘌呤(IMP)均未显著改变cAMP水平(图1F)。相反,嘌呤肌苷诱导细胞内cAMP显著增加(图1F, 图S1K),这表明肌苷可能在BAT激活和代谢中发挥重要作用。
为了进一步明确BAT中肌苷的来源,作者还检测了脂肪组织中的内皮细胞和成纤维细胞中的肌苷水平,发现UV照射后内皮细胞外肌苷浓度显著升高,而成纤维细胞外肌苷水平没有显著变化(图S1L)。有趣的是,与棕色脂肪细胞相比,这两种细胞类型胞外肌苷浓度均较低(图S1L,图1D)。这些结果表明,在所研究的细胞类型中,棕色脂肪细胞是BAT细胞外肌苷的主要来源。接下来,作者分析了嘌呤能酶(参与ATP降解级联反应的所有主要酶)的表达,包括外核苷三磷酸二磷酸水解酶(Ectonucleoside triphosphate diphosphohydrolase1, ENTPD1)和外5′-核苷酸酶(Ecto-5’-nucleotidase, NT5E),以及催化腺苷转化为肌苷的酶,即腺苷脱氨酶(Adenosine deaminase, ADA),发现这些酶在小鼠棕色脂肪细胞中的表达量均比白色脂肪细胞高(图S1M)。
拓展阅读
肌苷的功能
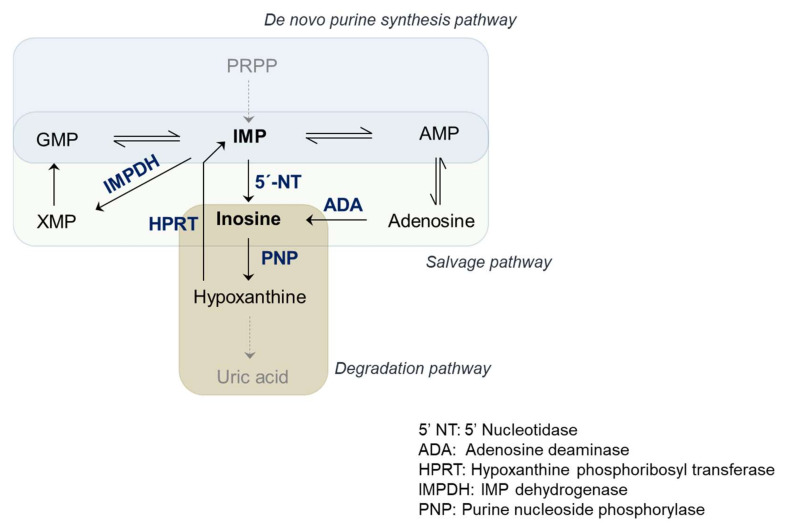
肌苷(Inosine),也被称为次黄苷,是由次黄嘌呤(Hypoxanthine)与核糖结合而成的核苷类化合物。由于其主要由肌肉中的ATP产物降解产生,因此也被称之为肌苷。另外,名字与肌苷类似的物质有肌酐和肌酸,它们是蛋白质代谢的产物,而肌苷是嘌呤代谢反应物,它们名字很像但并没有直接联系。在体内,肌苷为腺嘌呤的前体,能够直接透过细胞膜,参与机体内ATP、辅酶A、DNA及RNA的合成,起到调控机体的物质代谢和能量代谢的作用。多余的肌苷在嘌呤的降解途径中被嘌呤核苷磷酸化酶(Purine nucleoside phosphorylase, PNP)降解为次黄嘌呤,并进一步被分解为尿酸从体内排出。
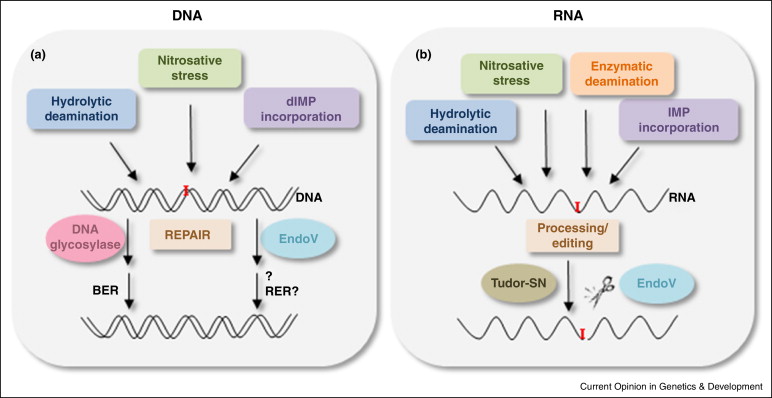
肌苷主要通过四种方式参与基因合成—腺苷自发水解、亚硝化诱导脱氨、IMP掺入和腺苷脱氨酶编辑(Adenosine deaminase action RNA, ADAR)。在RNA中,肌苷起两个主要的功能。在tRNA中,由于肌苷所引起的摆动作用,其可以与C-、A-或U-碱基结合,从而拓展了tRNA可以识别的三联体库,提高其翻译效率;在真核生物中,腺苷到肌苷(A到I)的编辑是常见的RNA修饰之一,由ADAR家族成员催化腺苷水解脱氨,如果在蛋白质编码区域发生修饰,则新产生的肌苷碱基在mRNA翻译过程中被核糖体解释为鸟苷,从而导致蛋白质产物改变。肌苷在mRNA中的掺入则可以改变双链区的三维结构,从而影响与RNA 结合蛋白的相互作用。因肌苷可以与腺苷在一定程度上相互转化,二者之间的动态调节也就起到了调控转录本定位,影响剪接和翻译准确性的功能。而在DNA中,脱氧肌苷由于其优先与胞嘧啶而非胸腺嘧啶结合的特性,会引起DNA位点发生突变,诱发DNA损伤。这些DNA可以被DNA糖基化酶识别,并通过碱基切除等方式进行修复。
参考文献:
[1] Guo W et al. J Agric Food Chem. 2021 Jul 14;69(27):7619-7628.
[2] Alseth I et al. Curr Opin Genet Dev. 2014 Jun;26:116-23.
[3] Srinivasan S et al. Genes (Basel). 2021 Apr 19;12(4):600.
3、肌苷通过cAMP/PKA轴发出信号
肌苷处理后细胞内cAMP显著增加,提示肌苷可能可以激活典型的cAMP-蛋白激酶A (PKA)通路。为了验证这一猜想,作者对使用肌苷或腺苷酸环化酶激活剂forskolin (forsk)处理的小鼠棕色脂肪细胞进行高灵敏磷酸化蛋白质组学分析,鉴定出38,451个磷酸肽。在肌苷或forsk处理的细胞中,分别有7875或8613个磷酸化位点被调控(FDR<0.05),其中2535个磷酸化位点在两种处理中均被显著调控 (图S2A)。有趣的是,通过数据库比对,作者发现PKA靶点在其中占据了非常大的部分(图S2A)。在forsk以及肌苷处理的棕色脂肪细胞中,p38丝裂原活化蛋白激酶(Mitogen-activated protein kinase, MAPK)轴激活并导致转录诱导激活转录因子2 (Activating transcription factor 2, ATF2)的Thr53和Thr51位点磷酸化(图1G)。此外,肌苷和forsk都下调了盐诱导激酶2 (Salt-inducible kinase 2, Sik2)的Ser576位点磷酸化水平,导致cAMP调节的转录共激活因子3 (cAMP-regulated transcriptional co-activator 3, Crtc3)下游位点去磷酸化(Ser72、Ser162、Ser329、Ser370)(图S2B)。这些位点的去磷酸化则可以诱导Crtc3的入核,并促进cAMP响应元件结合蛋白(cAMP-responsive element binding protein 1, Creb1)的结合和激活。此外,forsk和肌苷均促进了mTORC1和MAPK/ ERK信号途径,进而促进棕色脂肪细胞分化和棕色化(图S2C-D)。这些结果表明,肌苷激活PKA信号通路,PKA的下游靶点p38、Sik2、Crtc3和Creb控制核心棕色/米色基因程序和产热能力。Western Blot分析也证实了小鼠棕色脂肪细胞中MAPK及其下游靶点ATF2的激活,以及肌苷对Creb的激活(图S2E-G)。
肌苷处理还显著增加了小鼠棕色脂肪细胞中Ucp1的表达(图1H),Pparg的表达也有所提高 (图S2H)。肌苷也诱导成熟白色脂肪细胞棕色化,产热基因Ucp1和Ppargc1a的表达显著增加(图1I,图S2I),而在肌苷处理早期白色脂肪细胞时,脂肪生成抑制基因Necdin的水平降低(图2J)。此外,急性肌苷处理离体小鼠BAT可显著增加耗氧量和脂解能力(图1J-K;图S2K)。接下来,作者探究了在棕色脂肪细胞中介导肌苷作用的细胞受体(小编注:在脊椎动物中,存在四种腺苷受体,分别称为腺苷A 1、A 2A、A 2B和A 3,其中,肌苷对于A 2A、A 2B为完全激动剂,对于另外两种则仅起到部分激动作用)。考虑到肌苷对cAMP和脂解的刺激作用,作者重点研究了GS偶联的嘌呤能P1受体。在加入A2A拮抗剂MSX-2或A2B拮抗剂PSB603后可通过抑制甘油的释放来显著降低肌苷诱导的脂解作用,并且这两种拮抗剂组合使用消除了小鼠BA中的肌苷作用(图S2L)。

图S2 | 肌苷刺激产热
鉴于肌苷对产热脂肪细胞和BAT离体组织的刺激作用,作者采用间接量热法研究肌苷对全身能量消耗(EE)的影响。注射100μg/kg肌苷后,小鼠耗氧量显著增加(图1L)。为了研究A2A和A2B是否在体内介导肌苷作用,作者使用了A2A-KO和A2B-KO小鼠。向WT、A2A-KO或A2B-KO小鼠注射100μg/kg肌苷,并使用代谢笼监测耗氧量。与空白对照组相比,在WT小鼠中注射肌苷导致耗氧量增加(p<0.05),而这一效应在A2A-KO或A2B-KO动物中被抑制(图2M,N)。这些数据表明,肌苷信号通过在产热脂肪细胞中GS-偶联P1受体A2A和A2B激活cAMP/PKA/p38通路,从而提高能量消耗。
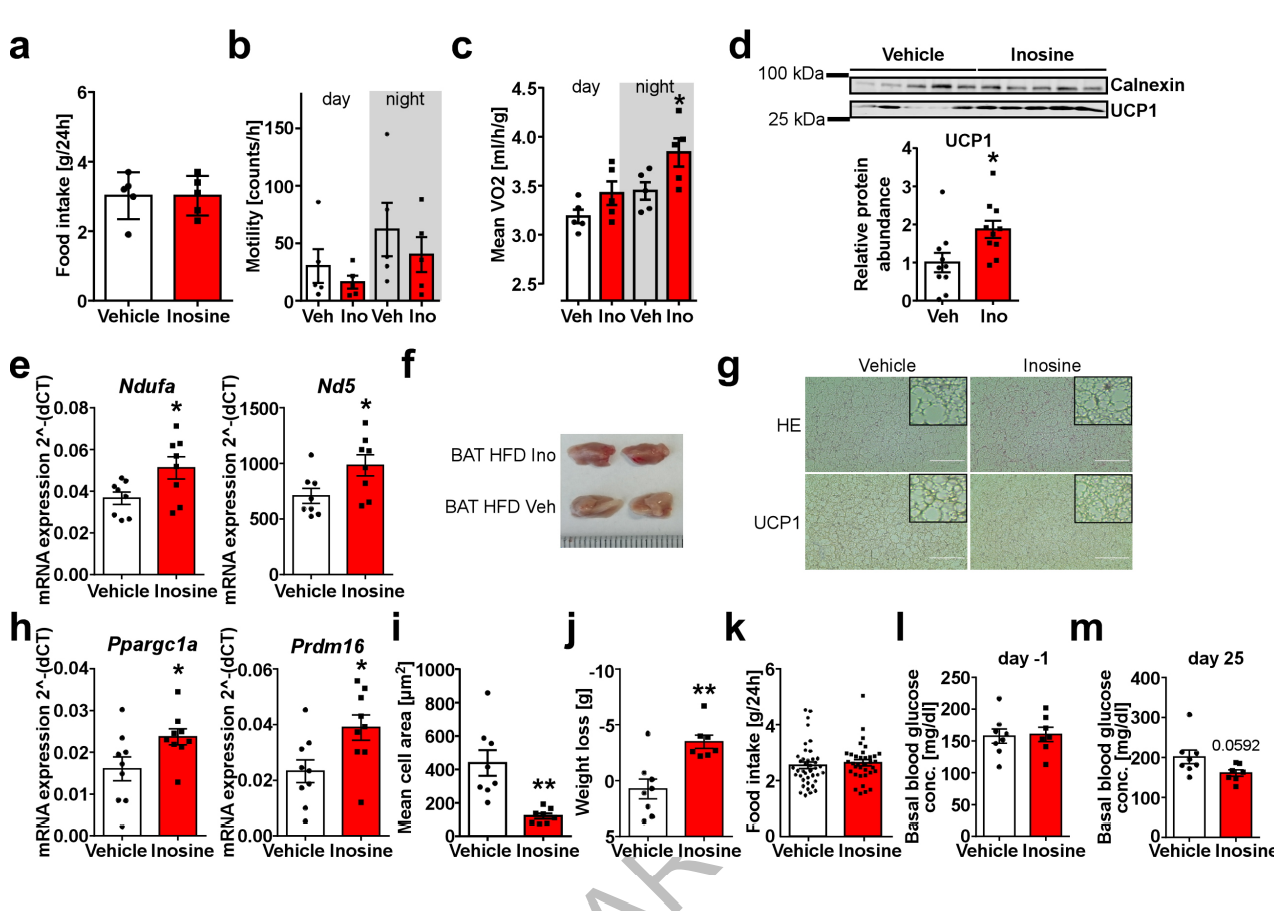
为了研究肌苷治疗对饮食诱导肥胖(DIO)是否有作用,作者在高脂肪饮食(HFD)的小鼠身上植入了微渗泵,并在四周内进行肌苷给药。结果表明,肌苷给药组的HFD小鼠体重显著低于对照组(图1M),且摄食量和运动能力没有受到影响(图S3A,B)。肌苷给药组小鼠耗氧量增加(图1N,图S3C),急性冷暴露(4℃)后棕色/米色脂肪产热能力显著增加(图1O),BAT中UCP1 mRNA和蛋白水平表达更高(图1P,图S3D),此外,肌苷给药组小鼠BAT中线粒体标记物(Ndufa和ND5)的表达上调 (图S3E),且BAT棕色化程度更深(图S3F)和脂滴更小(图S3G)。与对照组相比,肌苷给药组小鼠腹股沟白色脂肪组织(iWAT)的分析显示,Ucp1、Ppargc1a和Prdm16的表达显著增加(图1Q,图S3H),且组织学分析显示,在肌苷处理的小鼠iWAT中有更多的多房细胞和细胞大小减小,同时UCP1染色增强(图1R,图S3I)。这些数据表明,肌苷是一种新的BAT介导EE的激活剂,并诱导iWAT棕色化从而对抗DIO。
为了探究肌苷的治疗潜力,饮食诱导的肥胖小鼠每天注射对照溶媒剂或肌苷,为期26天,同时饲喂高脂饮食。从第7天起,与对照组相比,注射肌苷的小鼠的体重显著减轻(图1S)。因此,肌苷治疗组小鼠总体重减轻程度更高(图S3J)。注射肌苷的小鼠的脂肪重量显著减少,而对照小鼠的体成分没有明显变化(图1T),且两者之间摄食量没有差异(图S3K)。开始给药的前一天,两组的空腹血糖水平没有明显差异 (图S3L)。但在25天后,给药组空腹血糖水平显著下降(p=0.0592) (图S3M)。综上所述,这些数据进一步表明,肌苷可以减轻肥胖,具有一定的肥胖治疗潜力。
5、ENT1调节肌苷水平
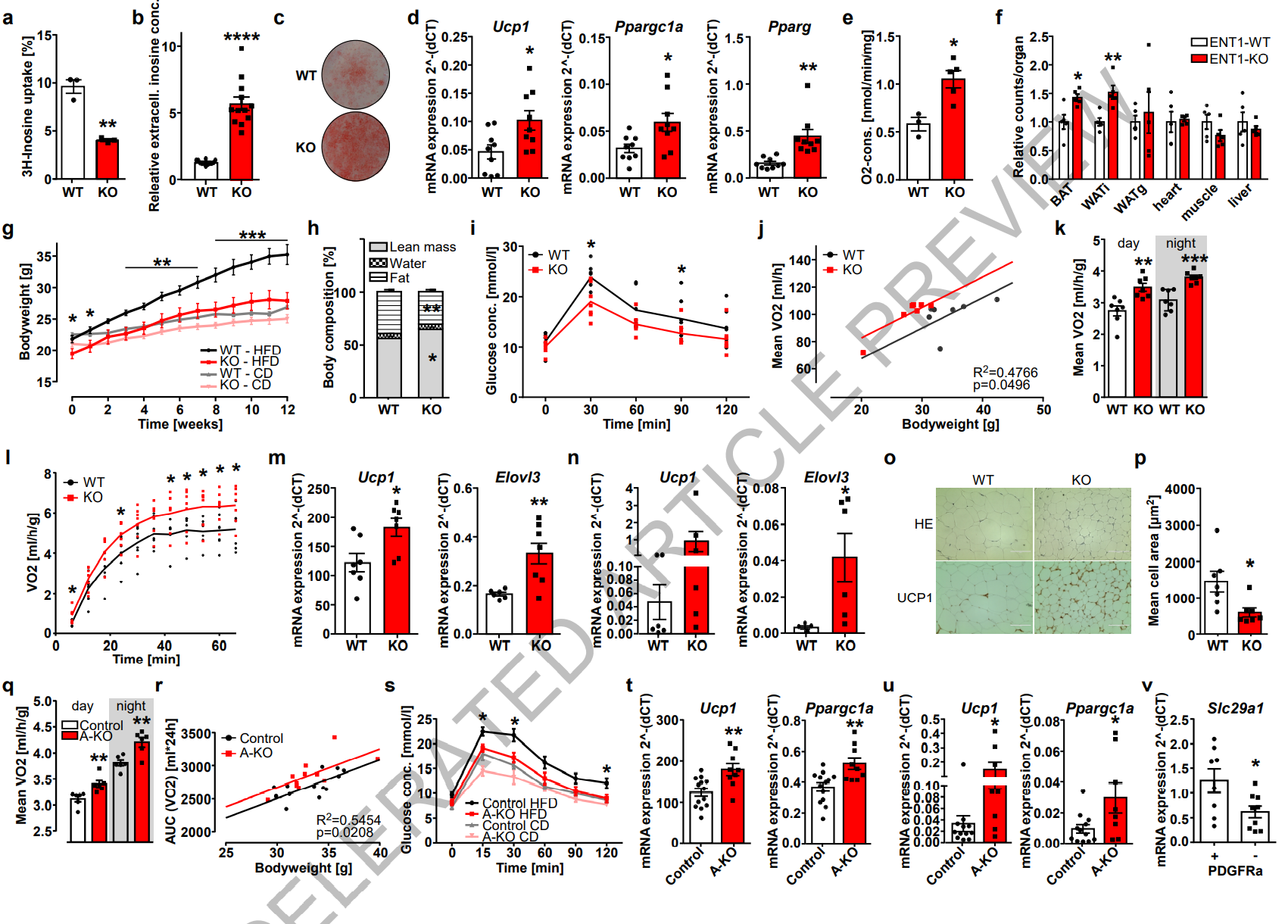
为了探究脂肪细胞如何调节胞外肌苷水平,作者重点研究了嘌呤在细胞膜上穿梭的转运体/通道。虽然Pannexin1通道在细胞凋亡中对核苷酸/ATP的释放十分重要,但ENT1和2也可以运输肌苷等核苷。其中,ENT1/SLc29a1最近被发现是棕色脂肪细胞的一个新的标记物。由于对ENTs在脂肪细胞中的功能知之甚少,本研究便分析了在分离的BAT、iWAT和性腺WAT(gWAT)原代脂肪细胞中编码ENT1和2的基因SLc29a1和Slc29a2的表达情况。SLc29a1是迄今为止在脂肪细胞中表达最高的ENT基因(图S4A),且棕色脂肪细胞表达ENT1的水平显著高于白色脂肪细胞(图S4A)。为了研究ENT1是否参与肌苷的穿梭,作者在ENT1缺陷(ENT1-/-)小鼠中分离出棕色脂肪细胞并测量3H标记肌苷的摄取。结果发现与WT对照细胞相比,ENT1-/-的棕色脂肪细胞从细胞培养上清液中摄入的3H-肌苷显著减少(图2A),在ENT1-/-棕色脂肪细胞的培养上清液中积累的肌苷更多(图2B)。对ENT1功能作用的分析表明,在ENT1-/-棕色脂肪细胞中,成脂和产热分化增强,脂滴形成增加(图2C),产热基因Ucp1和Ppargc1a以及成脂基因Adipoq、Fabp4和Pparg的表达升高(图2D,图S4B)。ENT1-/-脂肪细胞分化的增强与凋亡上清液和/或肌苷处理的效果一致(图1E,F)。此外,与WT细胞相比,ENT1-/-棕色脂肪细胞的耗氧量和脂解作用显著增加(图S4C-D)。对离体组织的分析显示,与WT的BAT相比,ENT1-/-的基础和UCP1介导的耗氧量显著增加(图2E,图S4E)。同样,ENT1-/-的BAT体外脂解率也显著高于WT组(图S4F)。这些数据表明,ENT1的敲除增加了肌苷浓度,从而导致棕色脂肪细胞和BAT激活,且分化和产热能力增强。

图S4. ENT1基因的全身敲除
拓展阅读
核苷转运蛋白
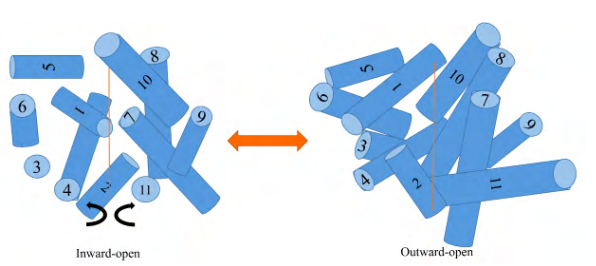
细胞外腺苷浓度由一组膜转运蛋白调节,这些蛋白都属于SLC超家族,特别是基因家族SLC28和SLC29。前者主要包括依赖于钠离子的集中核苷转运蛋白(Concentrative nucleoside transporter proteins, CNTs)和不依赖钠离子的平衡核苷转运蛋白(Equilibrative nucleoside transporter proteins, ENTs)。前者往往需要在高钠离子梯度中以1:1的比例摄取腺苷进入细胞从而参与基因或ATP等人体必需生理物质的生物合成;而后者则通过被动扩散摄取或排出腺苷,从而动态调节胞内外腺苷浓度。ENTs由11个疏水的跨膜结构域组成,通过不断变换为内向型构象和外向型构象从而起到转运功能。
ENTs主要包括1-4四种亚型。其中,ENT1广泛存在于所有细胞和组织中,是最主要的腺苷调节蛋白,ENT2在脑、心脏、胰脏和肾、骨骼肌等组织器官高表达。而ENT3和4则分别主要集中在胎盘组织和心脏、脑中发挥特异性功能。作为体内重要的腺苷平衡蛋白,ENT1主要通过调节腺苷、肌苷等核苷的浓度,影响细胞表面的受体激活水平,从而进一步调节信号传导。有研究报道表明,ENT1功能缺失引起的细胞外腺苷浓度上调可以导致肾纤维化和肾上皮细胞的上皮-间质转化(EMT),从而诱发糖尿病等一系列慢性肾病。但细胞外高浓度的腺苷也并非全部有害,通过敲除ENT1,研究发现心脏中较高浓度的细胞外腺苷可以激活腺苷受体起到保护心脏和神经、减轻炎症损伤、增强谷胱甘肽等ROS清除剂活性的作用。另外,ENT1主要介导摄取腺苷后参与生物合成,但是也有报道称ATP等激活物也可以介导其排出腺苷,激活细胞外腺苷受体,从而起到神经信号传导的功能。
参考文献: [1] Pastor-Anglada M et al. Front Pharmacol. 2018 Jun 14;9:627. [2] Baldwin SA et al. Pflugers Arch. 2004 Feb;447(5):735-43.
6、ENT1调节能量消耗
为了分析体内组织的能量摄取,研究人员对ENT1-/-和WT小鼠灌胃给药了放射性标记葡萄糖(3H-DOG)和脂肪酸(14C-三油酸脂)。在ENT1-/-小鼠中,BAT和WAT对14C-三油酸脂的摄取上调(图2F)。此外,iWAT的葡萄糖摄取能力也显著增加(图S4G)。两组小鼠的粪便中能量含量没有显著差异(小编注:推测是由于作者在给药2h后便进行处死,粪便获取时间偏早,因此粪便没有及时产生对应变化)(图S4H)。ENT1-/-白色脂肪细胞的成脂作用增强(小编注:作者发现iWAT中成脂相关基因和产热相关基因均显著上升,以此来说明其在外界高能环境中能量摄入和消耗均增强),表现为更多的脂滴以及成脂标记基因(ADIPOQ、FABP4和PPARG)表达增加(图S4I, J)。在ENT1-/-白色脂肪细胞中,产热基因Ucp1和Ppargc1a的表达显著增加,即白色脂肪细胞的棕色化程度增强(图S4J)。因此,与WT细胞相比,ENT1-/-白色脂肪细胞的基础和NE诱导的脂解作用和非偶联的呼吸作用增强(图S4K, L)。对iWAT外植体的分析显示,ENT1-/-的iWAT脂解能力和UCP1介导的呼吸显著增加(图S4M, N),基础呼吸增加了49%(但并不显著) (图S4N)。这些数据表明,ENT1在产热脂肪组织的能量消耗和WAT棕色化中起主要作用。
接下来,作者还探究了ENT1缺乏对DIO的影响。与WT小鼠比较,ENT1-/-小鼠在HFD条件下体重显著下降(图2G,图S4O),其脂肪重量也显著减少(图2H),iWAT和gWAT占比分别减少了26%和41%(图S4P)。此外,ENT1-/-小鼠在HFD后也表现出糖耐量改善(图2I,图S4Q)。与WT小鼠相比,ENT1-/-小鼠能量消耗增加(图2J, K, 图S4R),夜间活动减少(图S4S),而摄食量没有差异(图S4T)。在冷暴露(4℃)时,ENT1-/-小鼠的总产热能力以及ENT1-/-BAT和iWAT中的产热标记基因的表达显著增加(图2L-N,图S4U,V)。组织学分析显示,在ENT1-/-iWAT中(图2P),UCP1的表达上调1(图2O),脂肪细胞大小显著减小。总之,ENT1-/-小鼠表现出BAT功能增强和iWAT棕色化,且抵抗高脂饮食诱导的肥胖。
8、脂肪特异性敲除ENT1基因
鉴于ENT1的泛表达模式,不同类型的细胞可能在不同程度上参与肌苷的摄取。因此,本研究通过将Slc29a1-Floxed(ENT1f1)小鼠与脂联素启动子-Cr小鼠(ApnCre)杂交,构建了脂肪组织特异性敲除(A-KO)小鼠模型,导致BAT和iWAT中ENT1分别下调48%和57%(图S5A)。在A-KO小鼠的原代棕色脂肪细胞和原代白色脂肪细胞以及BAT和iWAT外植体中,成脂基因(Fabp4,Pparg,Adipoq)和产热基因(Ucp1, Ppargc1a)表达显著升高(图S5B,C),能量消耗(图S5D-F)和脂解(图S5G)作用增强。与ENT1fl小鼠相比,ENT1-A-KO小鼠的全身EE显著增加(图2Q,图S5H)。摄食量和通过粪便排出的能量没有显著变化(图S5I, J),而ENT1-A-KO动物的运动量较少(图S5K)。与ENT1-/-小鼠类似,在ENT1-A-KO的BAT中产热基因 (Ucp1、Ppargc1a、Prdm16)的表达显著增加(图S5L)。UCP1和Ppargc1a在ENT1-A-KO iWAT中的表达也增加(Ucp1上调得不够显著)(图S5M)。
在HFD的12周内,ENT1-A-KO小鼠与对照同窝小鼠相比体重减轻了13% (图S5N),并表现出了更高的EE/耗氧量(图2R)和糖耐量的显著改善(图2S),以及BAT和iWAT组织中产热基因Ucp1和Ppargc1a表达增加(图2T, U)。活动量、摄食量和肠道能量吸收在两种基因型之间没有明显变化(图S5O-Q)。为了进一步研究ENT1在脂肪细胞棕色化中的作用,作者重点研究了表达PDGFRα的前体细胞,这些细胞已被证明可分化为米色/棕色脂肪细胞。值得注意的是,与PDGFRα阴性的细胞相比,在PDGFRα阳性的iWAT间质血管部分细胞中的Slc29a1表达显著上调(图2V)。综上所述,脂肪组织特异性ENT1基因敲除可以重现ENT1-/-小鼠的表型,这表明脂肪细胞中ENT1在调节产热和全身EE中发挥着主要作用。

图S5. 脂肪细胞特异性敲除ENT1基因
9、ENTs的药理抑制作用
使用双嘧达莫(一种被批准的阻断ENTs的抗血小板药物)处理棕色脂肪细胞,导致细胞外肌苷显著增加(图S6A)并抑制肌苷摄取(图S6B)。尽管双嘧达莫单独给药可以诱导脂解增加 (图S6C),但这一效果与对照组相比没有显著差异,这提示必须先触发肌苷释放从而获得显著的ENT1药理抑制作用。为了研究除细胞凋亡外,还有哪些刺激可以诱导BAT内肌苷的积聚,作者推测肌苷可能在BAT的生理交感兴奋过程中积聚,这已被证明可以诱导肌苷前体腺苷的释放。事实上,使用去甲肾上腺素(NE)激活棕色脂肪细胞后,细胞外肌苷显著增加(图S6D)。为了直接分析BAT中的肌苷水平,作者建立了一个微透析装置,并发现在NE的急性激活或小鼠在冷暴露(4℃)7天后,肌苷水平分别增加了2.7倍和3.0倍(图S6E)。这些数据表明,BAT在生理刺激后也会释放肌苷,从而构成了一个迄今未知的正前馈环路,可能有助于在交感神经激活能量消耗后持续激活BAT。
拓展阅读
嘌呤、腺苷和肌苷
嘌呤核苷酸是指嘌呤与核糖、磷酸共同结合形成的化合物,在体内充当能量来源、代谢酶的辅助因子和信号分子。嘌呤核苷酸主要包括腺苷酸和鸟苷酸两种,其来源主要来自嘌呤从头合成途径,而肌苷酸(Inosine monophosphate,IMP)则作为嘌呤生物合成和降解途径的中心中间体在体内(尤其是肌肉中)广泛存在。在5-磷酸核糖与ATP在磷酸核糖焦磷酸合成酶催化下形成磷酸核糖焦磷酸 (PRPP) 后,PRPP进入嘌呤合成步骤,经10步的核糖修饰生成IMP,并在与天冬氨酸结合后裂解形成AMP和GMP,或通过5’-核苷酸酶催化产生肌苷。除上述途径外,肌苷也可以由AMP在腺苷脱氨酶(Adenosine deaminase, ADA)经脱氨反应获得。而在体内嘌呤含量较低时,嘌呤补救合成途径被激活,肌苷及其代谢物次黄嘌呤(Hypoxanthine)被次黄嘌呤磷酸核糖基转移酶(Hypoxanthine phosphoribosyltransferase, HPRT)还原为IMP,并在肌苷一磷酸脱氢酶(Inosine monophosphate dehydrogenase, IMPDH) 的催化下转变为GMP的前体单磷酸黄嘌呤 (Xanthine monophosphate, XMP)。从而在一定程度上补充体内的嘌呤缺乏。
嘌呤核苷腺苷是ATP的主要前体和代谢产物,它在交感神经系统中充当协同神经递质,其作用与腺苷的细胞外浓度和与腺苷受体的结合程度有关。腺苷信号主要和四种腺苷受体结合。在白色脂肪中,腺苷主要通过抑制型G蛋白Gi结合腺苷A1和A3受体,起到抑制脂解的作用;而在棕色脂肪中则恰好相反,通过激活型G蛋白Gs传递结合A2A和A2B受体,从而促进脂解的发生。
参考文献: [1] Srinivasan S et al. Genes (Basel). 2021 Apr 19;12(4):600. [2] Silva L et al. Mol Aspects Med. 2017 Jun;55:45-61. [3] Reverte-Salisa L et al. Handb Exp Pharmacol. 2019;251:161-182.
由于双嘧达莫与NE一起处理的棕色脂肪细胞对NE诱导的激活(脂分解)表现出显著的增加效应(图S6C),接下来作者又研究了在存在或不存在β3-肾上腺素受体激动剂CL-316 243(CL)的情况下注射双嘧达莫的小鼠,结果发现,使用浓度(0.3mg/kg)的CL仅导致轻微的、不显著的EE诱导(图S6F)。虽然双嘧达莫单独使用会导致小鼠耗氧量增加,但与对照组相比,双嘧达莫和CL共同使用会导致小鼠耗氧量进一步显著增加(图S6F)。同样,在冷暴露的小鼠中急性注射双嘧达莫可诱导氧消耗显著增加,这与在ENT1-/-小鼠中观察到的水平相当(图S6G)。
为了研究ENT1阻断是否对棕色化有影响,作者在WT小鼠冷暴露(4℃)期间注射双嘧达莫7天。与对照小鼠相比,双嘧达莫治疗导致小鼠的耗氧量显著增加(图S6H)。有趣的是,注射了双嘧达莫的小鼠与对照小鼠相比,BAT重量显著增加(图S6I),而iWAT和gWAT的组织重量分别减少了19%和10%(图S6I)。免疫组织化学分析显示,在双嘧达莫处理的小鼠iWAT中,脂肪细胞表面积较小,UCP1阳性细胞较多(图S6J)。这些数据表明,抑制ENTs协同增加了生理或药物激活BAT的EE,并增强iWAT的棕色化。
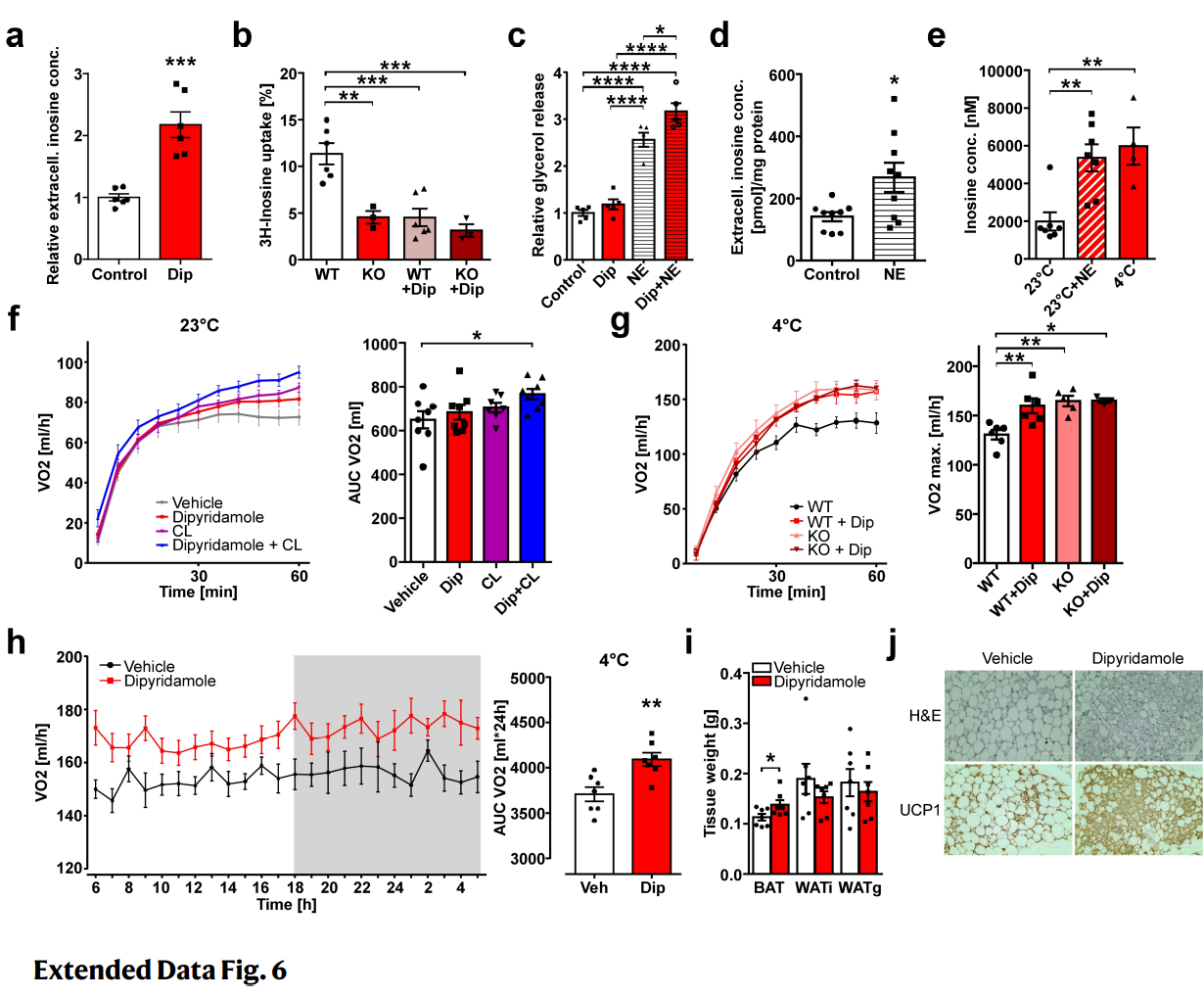
图S6. 双嘧达莫抑制ENT1对脂肪组织的影响
为了研究肌苷在人类棕色脂肪细胞(hBA)中的作用,作者从颈部活检组织中分离出人原代前体脂肪细胞,并将其分化为成熟的棕色脂肪细胞。在不破坏细胞膜的情况下,用Nutlin-3或紫外线处理诱导细胞凋亡。非靶向代谢组学显示,在两种凋亡条件下(Nutlin-3和紫外线照射),细胞外肌苷浓度都上调 (图S7A, B)。对嘌呤能分子的靶向分析进一步显示,Nutlin-3或紫外线照射处理后,hBA胞外肌苷浓度显著增加,且肌苷是释放最丰富的嘌呤(图3A,图S7C)。此外,用去甲肾上腺素刺激hBA可显著增加细胞外肌苷的积聚(图S7D)。
与小鼠棕色脂肪细胞相似(图1H),在hBA中用肌苷(300 nm)处理可以导致产热基因UCP1和TFAM以及成脂分化标记PPARG和FABP4的表达增加(图3B)。作者使用了Crispr-Cas9慢病毒载体敲除hBA中SLC29A1/ENT1;并筛选出效率最高的CRISPR3用以进行后续的ENT1敲除 (ENT1-KD) (图S7E)。与对照细胞相比,ENT1-KD脂肪细胞摄取3H-肌苷能力显著降低(图3C),这与双嘧达莫在hBA中的作用相似。这些发现与在双嘧达莫治疗和ENT1-Crispr转导的hBA中检测的细胞外肌苷浓度的增加是一致的(图S7F)。与小鼠ENT1-/-棕色脂肪细胞相似,成熟的ENT1-KD hBA显示PPARG和PRDM16的表达显著上调(图S7G),产热标记基因TFAM、UCP1和PPARGC1A的表达则分别增加22%、29%和15%(无显著差异)(图S7G)。
接下来,作者比较了人类棕色脂肪细胞和白色脂肪细胞中嘌呤能酶的表达水平,发现ATP降解酶ENTPD1和黄嘌呤氧化酶(Xanthine oxidase, XO)在hBA中的表达较高(图S7H)。ADA(将腺苷转化为肌苷的酶)在hBA中的表达也显著上调 (图S7H)。
为了研究肌苷对人白色脂肪细胞(hWA, human white adipocytes)和米色脂肪细胞的作用,作者分别使用了原代hWA和hMADS(小编注:分别为人类白色脂肪细胞和人类多能脂肪干细胞,分别用以代表肌苷在人体WAT和米色脂肪中的作用)细胞系。在hWA和hMADS中,肌苷可以显著增强脂解作用(图3D)。值得注意的是,在hWA中敲除ENT1导致Ucp1表达和耗氧量增加,这表明降低ENT1功能之后,hWA发生“棕色化”(图3E, F;图S7I)。综上所述,凋亡的hBA释放肌苷,抑制或敲除ENT1增加细胞外肌苷水平。肌苷/ENT1的调节对人棕色和米色脂肪细胞的分化和功能有重要影响。
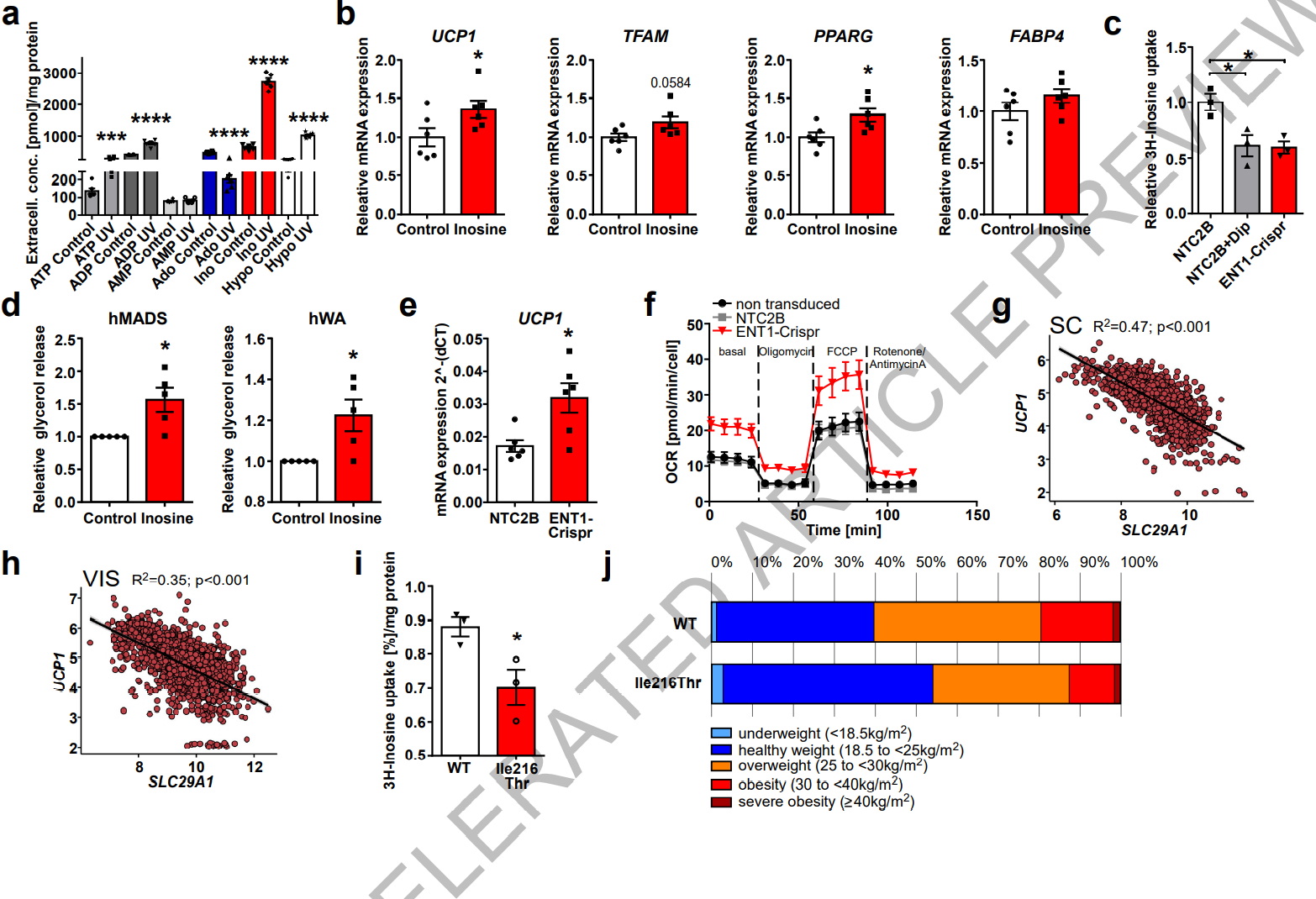
图S7. 肌苷和ENT1对人类脂肪细胞的影响
接下来,作者分析了人皮下和内脏WAT中的SLC29A1的表达与产热标记基因如UCP1、PGC1a和PRDM16表达的相关性(1400多名受试者)。结果发现,SLC29A1/ENT1的表达与UCP1 (图3G, H)、PPARGC1A和PRDM16(图S7J, K)表达呈显著负相关。这些数据表明,SLC29A1表达降低引起的细胞外肌苷浓度的增加,与人类WAT中产热基因的高表达成正相关。
对基因组聚合数据库(GnomAD)的分析发现,SLC29A1中存在多个具有错义突变的变体。在欧洲和非芬兰人群中频率最高的变异是SLC29A1的Ile216Thr突变(即第216位氨基酸由异亮氨酸突变苏氨酸)。重要的是,与WT变体的过表达相比,慢病毒载体过表达Ile216Thr变体导致3H-肌苷摄取显著减少(图3I)。因此,确定Ile216Thr突变为功能丧失突变。
为了解决Ile216Thr变异的潜在临床相关性,作者在一项广泛的表型研究队列中探究了Ile216Thr变异与BMI的相关性,该队列来自德国一个独立的SORB人群。根据排除标准(去除年龄在18岁以下、怀孕或哺乳期、急性感染和糖尿病的人群数据)将895名具有可用基因型的受试者进行分析。发现次要等位基因频率为4%,分布符合Hardy-Weinberg平衡(p>0.05)。值得注意的是,次要C等位基因与较低的平均体重指数显著相关(图3J)。最重要的是,在Thr变异携带者(Ile/Thr和Thr/Thr)组中,体重偏轻或健康的比例高于Ile/Ile纯合子受试者(55%对40%)(图3J)。与此相一致的是,与非携带者相比,Thr变异携带者肥胖(BMI<25 vs BMI>30 kg/m2)的几率降低了59%(图3J)。
总结
本研究发现,凋亡的棕色脂肪细胞通过释放肌苷增强了健康脂肪细胞产热程序的表达。该通路通过cAMP/蛋白激酶A信号通路刺激棕色脂肪细胞的能量消耗。利用肌苷治疗小鼠能够增加BAT依赖的能量消耗,并诱导白色脂肪组织“棕色化”。就机制而言,平衡核苷转运体1(ENT1)能够促进脂肪细胞对肌苷的摄取,降低细胞外肌苷水平,从而抑制脂肪组织的产热基因表达。而对ENT1的药理抑制以及敲除能够增强BAT活性并抵抗饮食诱导的肥胖。最后,人类ENT1的Ile216Thr功能丧失突变与BMI明显降低有关。
关注微信公众号代谢学人
了解更多代谢前沿资讯

https://m.sciencenet.cn/blog-3483272-1349007.html
上一篇:代谢学人--Nature Metabolism 6月刊代谢精选
下一篇:代谢学人--Science:中暑为啥会死人?