ВЉЮФ
ЗЪХжШнвзздЩэУтвпадМВВЁЕФдвђ
 ОЋбЁ
ОЋбЁ
|
ЗЪХжКЭздЩэУтвпМВВЁ
здЩэУтвпМВВЁЪЧжИЛњЬхЖдздЩэПЙдЗЂЩњУтвпЗДгІЖјЕМжТздЩэзщжЏЫ№КІЕФМВВЁЁЃздДгDonath гыLandsteinerЬсГіДЫИХФювдРДЃЌаэЖрМВВЁЯрМЬБЛСаЮЊздЩэУтвпадМВВЁЃЌГЃМћЕФШчРрЗчЪЊадЙиНкбзЁЂИЩдязлКЯжЂЁЂКьАпРЧДЏЁЂЦЄМЁбзЁЂМзПКЁЂЬЧФђВЁЁЂНсГІбзЕШЁЃжЕЕУЬсГіЕФЪЧЃЌздЩэПЙЬхЕФДцдкгыздЩэУтвпадМВВЁВЂЗЧСНИіЕШЭЌЕФИХФюЃЌздЩэПЙЬхПЩДцдкгкЮоздЩэУтвпадМВВЁЕФе§ГЃШЫЬиБ№ЪЧРЯФъШЫЃЌШчПЙМззДЯйЧђЕААзЁЂМззДЯйЩЯЦЄЯИАћЁЂЮИБкЯИАћЁЂЯИАћКЫDNAПЙЬхЕШЁЃгаЪБЃЌЪмЫ№ЛђПЙдадЗЂЩњБфЛЏЕФзщжЏПЩМЄЗЂздЩэПЙЬхЕФВњЩњЃЌШчаФМЁШБбЊЪБЃЌЛЕЫРЕФаФМЁПЩЕМжТПЙаФМЁздЩэПЙЬхаЮГЩЃЌЕЋДЫПЙЬхВЂЮожТВЁзїгУЃЌЪЧвЛжжМЬЗЂадУтвпЗДгІЁЃвђДЫЃЌвЊШЗЖЈздЩэУтвпадМВВЁЕФДцдквЛАуашвЊИљОнЃКЂйгаздЩэУтвпЗДгІЕФДцдкЃЌЂкХХГ§МЬЗЂадУтвпЗДгІжЎПЩФмЃЌЂлХХГ§ЦфЫћВЁвђЕФДцдкЁЃ
СюШЫаХЗўЕФСїааВЁбЇжЄОнБэУїЃЌГЌжиЛђЗЪХжгыЗЂЩњздЩэУтвпадМВВЁЕФЗчЯежЎМфДцдкКмЧПЕФЙиСЊЃЈ1ЃЉЁЃДгУтвпбЇЕФНЧЖШРДПДЃЌгыетжжЙиСЊЯрЙиЕФЯИАћКЭЗжзгЛњжЦАќРЈгЊбјКЭФмСПИагІЭООЖЖдTСмАЭЯИАћЕФЙ§ЖШДЬМЄЁЃИіЬхЕФУтвпДњаЛзДЬЌЪЧЕїНкУтвпздЮвФЭЪмадЕФКЫаФЃЌУтвпФЭЪмадвжжЦздЮвЗДгІвдБмУтздЩэУтвпЁЃжЌЗОзщжЏЪЧвЛжжУтвпЛюадЦїЙйЃЌЭЈЙ§ВњЩњжЌЗОЯИАћвђзггАЯьШЋЩэУтвпЗДгІЃЌЗДЙ§РДЃЌУтвпЯИАћЭЈЙ§ВњЩњДйбзКЭПЙбзЯИАћвђзггАЯьжЌЗОЯИАћЮШЬЌКЭДњаЛЃЈ2ЃЉЁЃетвтЮЖзХЗЪХжв§Ц№ЕФДњаЛГЌИККЩЛсгАЯьУтвпДњаЛЃЌДгЖјИФБфЖдздЩэУтвпадМВВЁЕФвзИаадЁЃ
The link between obesity and autoimmunity | Science
УтвпЪЪгІЪЧЖдгЊбјзДПіЕФЗДгІЃКгЊбјВЛСМЛсЫ№КІУтвпСІЃЌЕМжТЖдИаШОКЭвпУчНгжжЕФЗДгІаЇТЪЕЭЯТЁЃЯрЗДЃЌгЊбјЙ§ЪЃгаРћгкЯШЬьадКЭЪЪгІадУтвпЯИАћЕФТ§адМЄЛюЃЌЫцКѓГіЯжЃЈЕЭЖШЃЉШЋЩэбзжЂЁЃетаЉЯжЯѓЭЈЙ§ЯИАћФкгЊбјКЭФмСПИагІЭООЖвдМАКЌгаЕААзжЪ3ЃЈNLRP3ЃЉЕФNACHTЃЌLRRКЭPYDНсЙЙгђбзжЂаЁЬхЕФВЮгыЖјЗЂЩњЃЌетЪЧгЩЙ§СПЕФЦЯЬбЬЧКЭжЌжЪгеЕМЕФДњаЛгІМЄЕФДЋИаЦїЃЌЬиБ№ЪЧдкОоЪЩЯИАћжаЃЈ2ЃЌ3ЃЉЁЃ
ЗЪХжЪЧздЩэУтвпадМВВЁЕФЮЃЯевђЫиЃЌШч1аЭЬЧФђВЁЃЈT1DЃЉКЭЖрЗЂадгВЛЏжЂЃЈMSЃЉЃЈ4ЃЌ5ЃЉЁЃдіМг ЖрЗЂадгВЛЏжЂЗчЯеЕФЛЗОГКЭЩњЛюЗНЪНвђЫиАќРЈЮќбЬЁЂШеЩЙЁЂЮЌЩњЫи D ЕЭЁЂEB ВЁЖОИаШОКЭИпЬхжижИЪ§ ЃЈBMIЃЉЁЃЖдФъЧсЗЪХжИіЬхЕФЧАеАадзнЯђбаОПЗЂЯжЃЌЧрДКЦкКЭГЩФъдчЦкЃЈЕЋВЛЪЧдк MS ЗЂВЁЪБЃЉЛМ ЖрЗЂадгВЛЏжЂЕФЗчЯедіМгСЫ 1.6 жС 1.9 БЖ;етжжгыЗЪХжЕФЙиСЊвВдкШЫАзЯИАћПЙдЃЈHLAЃЉЈCDRB1*15:01взИаЕШЮЛЛљвђЕФаЏДјепжаЕУЕНжЄЪЕЃЌИУЕШЮЛЛљвђИКд№НЋЫшЧЪздЩэПЙдГЪЕнИјздЩэЗДгІадTЯИАћЃЈ5ЃЉЁЃЭЌбљЃЌГіЩњЪБНЯИпЕФBMIгыЖљЭЏНЯИпЕФT1DвзИаадгаЙиЁЃЪТЪЕЩЯЃЌT1DЕФЗЂВЁТЪМИКѕЫцзХГіЩњЬхжиЕФдіМгЖјЯпаддіМгЃЈГіЩњЬхжиУПдіМг100gЃЌЗЂВЁТЪдіМг1.7%ЃЉЃЈ4ЃЉЁЃ
ДгЛњжЦЩЯНВЃЌгаШЫШЯЮЊЃЌЩэЬхЗЪХжЕФдіМгЛсДйНјЯИАћФкгЊбјКЭФмСПИагІЭООЖ[Р§ШчРзХСУЙЫиЕФЛњжЦАаБъЃЈmTORЃЉ]ЕФЙ§ЖШМЄЛюЃЌЫцКѓЭтжмзщжЏжаЕФДњаЛГЌИККЩЃЌАќРЈВЮгыаЇгІКЭЕїНкУтвпЗДгІЕФГЃзЄУтвпЯИАћЃЈ6).Р§ШчЃЌдкЗЪХжЕФГѕжЮЖрЗЂадгВЛЏжЂЛМепжаЃЌЗЂЯжжЌЗОЯИАћвђзгЪнЫиЃЈгыBMIГЩБШР§ЗжУквдвжжЦЪГЮяЩуШыЃЉвдМАЩ§ИпЕФбЛЗгЊбјЫиПЩвддіЧПбзжЂУтвпЗДгІЁЃИпЫЎЦНЕФЪнЫиКЭгЊбјЮяжЪЕМжТTЯИАћжаmTORЕФзщГЩадЙ§ЖШМЄЛюЃЌЫцКѓTЯИАћЪмЬхЃЈTCRЃЉНщЕМЕФаХКХДЋЕМЪЇЕїЁЃTЯИАћжаЙ§ЖШЛюдОЕФmTORФЃЗТЧПСвЕФГЌЩњРэTCRДЬМЄЃЌетжжДЬМЄВЛдЪаэзЊТМВцЭЗКаP3ЃЈFOXP3ЃЉЛљвђЃЌЦфБэДяЖдгкгеЕМКЭЮЌГжПЙбзCD4CD25FOXP3ЕїНкадTЯИАћЃЈT+++зЂВс) (2ЃЌ6ЃЉЁЃЭЈЙ§ЪнЫиЙ§ЖШЩњВњЃЌЗЪХжЛсЫ№КІПЙбзаиЯй T ЕФдіжГзЂВсМАЦфгыCD4CD25ЕФЭтжмЗжЛЏ+−ДЋЭГT ЃЈTconvЃЉ ЯИАћЧАЬх ЃЈ7ЃЉЁЃЗЪХжвВЛсДйНј T ЕФзЊЛЏДЋЭГЯИАћзЊЛЏЮЊжТВЁадбзжЂадTИЈжњадTЃЈTH1ЃЉ КЭ TH17ИіЯИАћЃЌДгЖјдіМгУтвпздЮвФЭЪмадИФБфЕФЗчЯеЃЈМћЭМЃЉЁЃ
змЬхЖјбдЃЌгЊбјКЭЪнЫигеЕМЕФmTORЙ§ЖШМЄЛювжжЦЭтжмTregдіжГКЭвжжЦЙІФмЃЌдіЧПЗЪХжЯрЙиЕФTH1 КЭ TH17ЯИАћЗжЛЏЃЌОпгаНЯИпЕФЖрЗЂадгВЛЏЯрЙиЫшЧЪЫ№ЩЫЗчЯеЃЈ2ЃЌ7ЃЉЁЃДЫЭтЃЌзюНќЕФвЛЗнБЈИцБэУїЃЌЗЪХжаЁЪѓзЊЛЛСЫОЕфЕФTH2-ЬигІадЦЄбзЕФУтвпЗДгІжївЊЮЊвд T ЮЊЬиеїЕФбЯжиМВВЁHгЩЙ§бѕЛЏЮяУИЬхдіжГЮяМЄЛюЕФЪмЬхІУЃЈPPAR-ІУЃЉзЊТМвђзгЛюадНЕЕЭв§Ц№ЕФ17Ч§ЖЏаЭбзжЂЃЈ8ЃЉЁЃPPAR--дкжЌЗОзщжЏжаЕФБэДяЖдгкжЌЗОзщжЏзЄСєTregЕФЗЂг§КЭЙІФмвВЪЧБиашЕФЁЃБэУїжЌЗОзщжЏЩњЮябЇгыУтвпФЭЪмжЎМфДцдкЩцМА TregЕФЫЋЯђСЊЯЕЃЌ (2ЃЌ7ЃЉЁЃ
гЩгкУПШеНћЪГКЭНјЪГжмЦкв§Ц№ЕФЩњРэгЊбјЫиКЭЪнЫиВЈЖЏОіЖЈСЫmTORЛюадЕФеёЕДЃЌетаЉеёЕДдкЗЪХжжавђЪГЮяЩуШыЙ§ЖрЖјЫ№ЪЇЁЃвђДЫЃЌдкОпгае§ГЃBMIКЭНјЪГКЭНћЪГЩњРэжмЦкЕФИіЬхжаЃЌздЮвФЭЪмадЕФЮЌГжКЭбгајгыTregжаmTORЛюадЕФеёЕДгаЙи.етЫЦКѕЪЧ TregЫљБиашЕФРЉдіКЭЙІФмзувдвжжЦжТВЁад TH1 КЭ TH17ЯИАћЃЌвђДЫздЩэУтвпЃЈ2ЃЌ6ЃЌ7ЃЉЁЃ
жЕЕУзЂвтЕФЪЧЃЌmTORЪЧАБЛљЫсЃЌЦЯЬбЬЧКЭжЌжЪДњаЛЪЎзжТЗПкЕФЙиМќЯИАћФкНкЕуЁЃДЫЭтЃЌгыгЊбјКЭДњаЛЯрЙиЕФЩњГЄвђзгЃЌШчЪнЫиЃЌвШЕКЫиКЭвШЕКЫибљЩњГЄвђзг1ЃЈIGF-1ЃЉЃЌМЄЛюУтвпЯИАћжаЕФmTORаХКХДЋЕМЃЌДгЖјгАЯьШЋЩэКЭЯИАћФкУтвпДњаЛЃЌДгЖјгАЯьбзжЂКЭздЩэУтвпЃЈ2ЃЌ6ЃЉЁЃжЌЗОзщжЏЛЙЗжУкбзадЯИАћвђзгЃЌШчАзЯИАћНщЫи-1 ЃЈIL-1ЃЉЁЂжзСіЛЕЫРвђзг-ІС ЃЈTNF-ІСЃЉЁЂIL-6ЁЂIL-17 КЭИЩШХЫи-ІУ ЃЈIFN-ІУЃЉЃЌвдМАЪнЫиЃЌЕМжТЖдЭтжмзщжЏЫ№ЩЫКЭздЩэУтвпЕФУєИаадИќИпЁЃвђДЫЃЌЮвЬсГіДњаЛЙЄзїСП - гЩгЊбјЫиЃЌжЌЗОЯИАћбмЩњЕФЩњГЄвђзгКЭжЌЗОЯИАћвђзггеЕМ - ПЩФмДњБэЭЈГЃЪГгУжТХжЮїЗНвћЪГЕФШЫЕФздЩэУтвпадМВВЁЕФМгЫйЦїЁЃ
дкаЁЪѓКЭШЫРржавбОжЄУїЃЌЪЪгІадКЭЯШЬьУтвпЯИАћПЩвджБНггАЯьЕМжТЗЪХжКЭЗЪХжЯрЙиДњаЛвьГЃЕФВЁРэЩњРэЪТМўЃЈ2ЃЉЁЃетвВПЩФмгажњгкМѕЩйTregдкЗЪХжШЫШКжаЙлВьЕНЕФЪ§зжЁЃжЌЗОзщжЏКЭУтвпЯЕЭГжЎМфДцдкНтЦЪбЇКЭЙІФмДЎШХЁЃЪТЪЕЩЯЃЌдЗЂадСмАЭЦїЙйЃЈЙЧЫшКЭаиЯйЃЉКЭДЮМЖСмАЭЦїЙйЃЈСмАЭНсЃЉЭЈГЃЧЖШыжЌЗОзщжЏжаВЂБЛжЌЗОзщжЏАќЮЇЁЃетжжСЌајаддЪаэTЯИАћЃЌTregЁЂBЯИАћЁЂЪїЭЛзДЯИАћКЭОоЪЩЯИАћЙщГВЕНжЌЗОзщжЏЁЃДЫЭтЃЌжЌЗОЯИАћПЩвдБэДяУтвпбљааЮЊЃЈ2ЃЉЁЃР§ШчЃЌжЌЗОЯИАћПЩвдЪЙгУгыЯШЬьУтвпЯИАћЯрЭЌЕФКЫНсКЯЙбОлЛЏНсЙЙгђ1ЃЈNOD1ЃЉВЁдЬхДЋИаЯЕЭГЧхГ§ЯИАћФкЯИОњЃЈ9ЃЉЁЃTreg ЕФБфЛЏдкЗЪХжжаЙлВьЕНЕФЪ§СПКЭЙІФмвВПЩФмгАЯьЖдИаШОКЭАЉжЂЕФвзИаадЃЈ2ЃЉЁЃЪТЪЕЩЯЃЌбЯжиМБадКєЮќзлКЯеїЙкзДВЁЖО2ЃЈSARS-CoV-2ЃЉИаШОгыздЩэПЙЬхЕФВњЩњгаЙиЃЌдкЗЪХжИіЬхжаИќЮЊбЯжиЃЈ10ЃЉЁЃДЫЭтЃЌЗЪХжШЫШКЕФАЉжЂУтвпжЮСЦЗДгІгХгкBMIНЯЕЭЕФЛМепЃЈ2ЃЉЁЃ
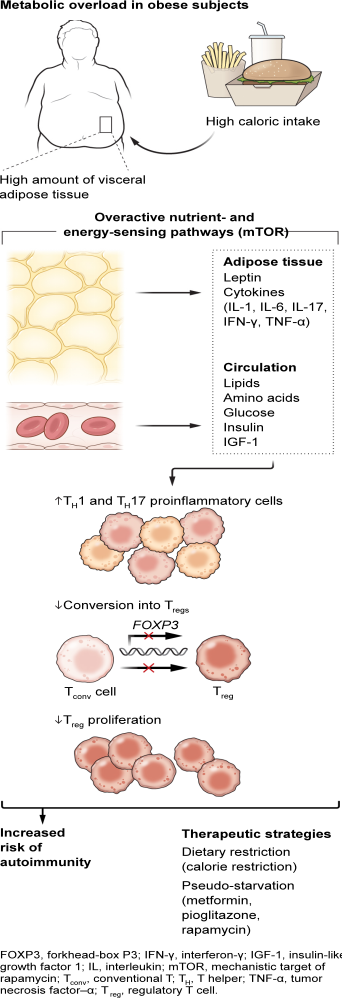
ЗЪХжжЂУтвпДњаЛИФБфПЩФмЕМжТздЩэУтвп
дкЗЪХжШЫШКжаЃЌЭЈЙ§бЛЗКЭжЌЗОзщжЏбмЩњвђзгЙ§ЖШДЬМЄУтвпЯИАћгЊбјКЭФмСПИагІЭООЖЃЈР§ШчдіМгmTORЛюадЃЉЛсХЄЧњУтвпЯИАћЗжЛЏЁЃетЛсдіМгДйбз T H1КЭ TH17ЯИАћЕФЪ§СПЃЌНЕЕЭTregЪ§СПЃЌетдіМгСЫЪЇШЅздЮвУтвпФЭЪмЕФЗчЯеЁЃУтвпДњаЛВпТдПЩФмгажњгкЛжИДЬхФкУтвпЦНКтЁЃ
ЖрЛљвђЗЪХжЃЈгЩЖржжвХДЋБфвьКЭЛЗОГвђЫив§Ц№ЕФЗЪХжвзИаадЃЉвВБЛШЯЮЊЪЧвЛжжздЩэУтвпбљМВВЁЃЌTЯИАћЖдЮДжЊЕФжЌЗОЯИАћПЙдзїГіЗДгІВЂДЅЗЂЫцКѓВЛЪмПижЦЕФЪГЮяЩуШыЃЌОЁЙмЦфЛњжЦШдгаД§ЭъШЋВћУїЃЈ11ЃЉЁЃжЕЕУзЂвтЕФЪЧЃЌДгЗЪХжаЁЪѓжаЗжРыЕФCD4 TЯИАћПЩвдЭЈЙ§НЋЬхжидіМгзЂЩфЕНе§ГЃЬхжиЃЌУтвпШБЯнЕФЪмЬхжаРДзЊвЦЁАЗЪХжМЧвфЁБЃЈ11ЃЉЁЃвђДЫЃЌЗЪХжЫЦКѕгыЗЂЩњздЩэУтвпЕФвзИаадИќИпгаЙиЃЌВЛНівђЮЊжЌЗОзщжЏЛсдіЧПздЩэбзжЂЗДгІЃЌЛЙвђЮЊЗЪХжБОЩэОпгаздЩэУтвпбљЬиеїЁЃ+
вЛжжгаЯЃЭћЕФПЩФмадЪЧЭЈЙ§УтвпДњаЛИЩдЄРДВйзнУтвпФЭЪмКЭздЩэУтвпЃКМѕЩйЪГЮяКЭ/ЛђПЈТЗРяЩуШыСПЁЃЫфШЛНћЪГПЩвдЕїНкУтвпЗДгІКЭЛКНтздЩэУтвпадМВВЁжЂзДЕФЯыЗЈДѓЖрБЛВЕЛиЃЌЕЋЙ§ШЅ20ФъЕФбаОПЬсЙЉСЫжЄОнЃЌжЇГжааЮЊИФБфКЭгЊбјВпТдЕФжЮСЦЧБСІЃЌШчвћЪГЃЌШШСПЯожЦЃЈCRЃЉКЭВЛЭЌЕФНћЪГЗНАИЃЈ2ЃЌ12).ЧсЖШШШСПЯожЦЃЌМфаЊадНћЪГКЭЩњЭЊвћЪГдкздЩэУтвпаЁЪѓФЃаЭжаОљЯдЪОГігавцЕФаЇЙћЃЌАќРЈЪЕбщадздЩэУтвпадФдМЙЫшбзЃЈEAEЃЉЃЌЪЕбщадРрЗчЪЊадЙиНкбзКЭЪЕбщадНсГІбзЃЈ2ЃЌ12ЃЉЁЃЮвНЈвщЁАМЂЖіЁБжТВЁадбзжЂTH1 КЭ TH17ИіЯИАћПЩвдИќКУЕиПижЦОжВПКЭШЋЩэбзжЂЁЃРрЫЦЕиЃЌШШСПЯожЦдЪаэTregЕФдіжГдкаЁЪѓКЭШЫРржаЭЈЙ§ДйНјЫќУЧЕФВњЩњЃЌдіжГКЭЙІФмЃЌДгЖјПижЦздЩэУтвпЃЈ2ЃЌ7ЃЌ12ЃЉЁЃ
гЩгкМсГжвћЪГИФБфВЂВЛзмЪЧПЩФмЕФЃЌвђДЫЬсГіЕФЬцДњЗНЗЈЪЧЁАМйМЂЖіЁБЃЌМДЕїНкУтвпДњаЛЕФвЉЮяФЃЗТНћЪГЃЈ13ЃЉЁЃвЛИіЕфаЭЕФР§згЪЧmTORвжжЦМСРзХСУЙЫиЁЃДЫЭтЃЌЖўМзЫЋывЪЧAMPЛюЛЏЕААзМЄУИЃЈAMPKЃЉЕФМЄЛюМСЃЌгУгкжЮСЦ2аЭЬЧФђВЁКЭГЌжиИіЬхЃЌВЛНіПЩвдПижЦЦЯЬбЬЧФЭСПЃЌЛЙПЩвдЭЈЙ§AMPKНщЕМЕФmTORвжжЦОпгаПЙбззїгУЃЈ13ЃЉЁЃЖўМзЫЋывЭЈЙ§ЯожЦЕЅКЫЯИАћНўШѓЕНжаЪрЩёОЯЕЭГ ЃЈCNSЃЉ ВЂЯТЕїбзжЂЯИАћвђзгЁЂгеЕМад NO КЯУИЁЂЯИАћеГИНЗжзгЁЂЛљжЪН№ЪєЕААзУИ-9 КЭЧїЛЏвђзгдк T жаЕФБэДяРДМѕШѕ EAE геЕМH17 ЯИАћ ЃЈ14ЃЉ.дкЖдЛМгаДњаЛзлКЯеїЕФЖрЗЂадгВЛЏЛМепЕФбаОПжавВЙлВьЕНСЫетаЉгАЯьЃЈ15ЃЉЁЃ
ЖрЗЂадгВЛЏжЂЃЈIFN-ІТЛђДзЫсИёРФЌЃЉЕФвЛЯпвЉЮяжЮСЦгыЖўМзЫЋывСЊКЯЪЙгУЃЌПЩЯджјИФЩЦМВВЁВЂМѕЩйжаЪрЩёОЯЕЭГВЁБфЁЃетаЉгАЯьгыбЛЗЪнЫиКЭTH1 КЭ TH17НЕЕЭЖржжбзадЯИАћвђзгКЭЭтжмTregЪ§СПдіМггаЙи(15ЃЉ. ЭЌбљЃЌОпгаПЙЬЧФђВЁзїгУЕФPPAR-ІУМЄЛюМСпСИёСаЭЊвВЭЈЙ§діМгвШЕКЫиУєИаадКЭНЕЕЭбЛЗЦЯЬбЬЧКЭЪнЫиЫЎЦНЃЌЮЊжЮСЦЖрЗЂадгВЛЏЛМепЕФУтвпЯИАћЬсЙЉМйМЂЖіЕФДњаЛаХКХЃЈ15ЃЉЁЃдкEAEжаЃЌпСИёСаЭЊжЮСЦПижЦСЫВЁГЬЃЌМѕЩйСЫCNSНўШѓЃЌМѕЩйСЫбзадЯИАћвђзгЕФВњЩњКЭTH1 КЭ TH17 ЗжЛЏ ЃЈ15ЃЉ.ДЫЭтЃЌгаШЄЕФЪЧЃЌОЕфЕФПЙбзКЭУтвпвжжЦвЉЮяШчЫЎбюЫсКЭМзАБЕћпЪПЩвдЭЈЙ§МЄЛюAMPKНЋМйМЂЖіЕФДњаЛаХКХДЋЕнИјУтвпЯИАћЃЈ13ЃЉЃЌвдМАЫќУЧЕФОЕфзїгУЛњжЦЁЃзмЬхЖјбдЃЌИГгшМйМЂЖіЕФДњаЛаХКХПЩФмЖдЯТЕїздЩэбзжЂЗДгІгаМлжЕЁЃ
жЕЕУзЂвтЕФЪЧЃЌдкШШСПЯожЦЦкМфЃЌTЯИАћНЋЦфзЊТМЬиеїжиаТБрГЬЮЊПЙбзЬиадЃЌЯожЦзщжЏЫ№ЩЫВЂбгГЄаЁЪѓКЭШЫРрЕФЪйУќЃЈ7ЃЌ12ЃЉЁЃДЫЭтЃЌШШСПЯожЦгеЕМГІЕРЮЂЩњЮяШКЙуЗКЪЪгІгАЯьОжВПКЭШЋЩэУтвпДњаЛЕФПЙбзДњаЛЮяЕФВњЩњЃЈ12ЃЉЁЃгыжЌЗОЯИАћбмЩњЕФЪнЫиЯрЛЅзїгУЕФЗжзгПЩвдИљОнДњаЛзДЬЌвдИїжжЗНЪНЕїНкУтвпЙІФмЁЃР§ШчЃЌОпгаЪГгћДЬМЄЛюадЕФЩёОФкЗжУкНщжЪЃЈШчЩњГЄЫиЪЭЗХыФКЭЩёОыФYЃЉгыЪнЫиЕФзїгУЯрЗДЃЌВЛНіЖдБЅИЙИаЖјЧвЖдЭтжмУтвпЗДгІЖМгаЯрЗДЕФзїгУЃЌвђЮЊЫќУЧЛсзшжЙTH1 КЭ TH17ЗжУкЖржжЯИАћвђзгВЂвжжЦEAEЃЈ2ЃЉЁЃЦфгрЕФбаОПСьгђАќРЈЕЅвЛгЊбјЫиЃЈМДжЌжЪЁЂЬМЫЎЛЏКЯЮяКЭЕААзжЪЃЉШчКЮгАЯьУтвпздЮвФЭЪмадЕФЗжзгНтЦЪЃЌвдМАШШСПЯожЦЪЧЗЪХжЯрЙиздЩэУтвпЕФгааЇжЮСЦЗНАИЕФЪБМфДАПкЁЃ
References
1
1. Lerner, P. Jeremias, T. Matthias, Int. J. Celiac. Dis 3, 151 (2015).
2. P. de Candia et al., J. Exp. Med. 3, 218 (2021).
3. Christ et al., Cell 172, 162 (2018).
4. A. March, D. J. Becker, I. M. Libman, Front. Endocrinol. 22, 12 (2021).
5. K. Hedström et al., Neurol. Neuroimmunol. Neuroinflamm. 8, e912 (2020).
6. M. H. Do et al., J. Exp. Med. 217, e20190848 (2020).
7. P. de Candia, C. Procaccini, C. Russo, M. T. Lepore, G. Matarese, Immunity 55, 1981 (2022).
8. S. P. Bapat et al., Nature 604, 337 (2022).
9. G. Caputa et al., Cell Metab. 34, 747 (2022).
10. E. Y. Wang et al., Nature 595, 283 (2021).
11. J. Zou et al., Cell. Mol. Immunol. 15, 630 (2018).
12. O. Spadaro et al., Science 375, 671 (2022).
13. L. A. OЁЏNeill, D. G. Hardie, Nature 493, 346 (2013).
14. Y. Sun et al., J. Neuroimmunol. 292, 58 (2016).
15. L. Negrotto, M. F. Farez, J. Correale, JAMA Neurol. 73, 520 (2016).
https://m.sciencenet.cn/blog-41174-1382744.html
ЩЯвЛЦЊЃКШчКЮзЋаДбЇЪѕТлЮФЃПЁЖПЦбЇЁЗЗУЬИ
ЯТвЛЦЊЃКасОѕеЯАВтЪдЕФеяЖЯМлжЕ