博文
[转载]348.个性化RNA新抗原疫苗刺激胰腺癌中的T细胞
|||
个性化RNA新抗原疫苗刺激胰腺癌中的T细胞
摘要
胰腺导管腺癌(PDAC)在88%的患者中是致命的,但含有适用于疫苗的突变衍生的T细胞新抗原。在佐剂自体cevumeran(一种基于尿苷mRNA-脂质体纳米颗粒的个体化新抗原疫苗)的I期试验中,我们从手术切除的PDAC肿瘤中实时合成了mRNA新抗原疫苗。手术后,我们依次给予阿替唑单抗(一种抗PD-L1免疫疗法)、自体cevumeran(每位患者最多20个新抗原)和一种改进的四药化疗方案(mFOLFIRINOX,包括亚叶酸、氟尿嘧啶、伊立替康和奥沙利铂)。终点包括通过高阈值检测的疫苗诱导的新抗原特异性T细胞、18个月无复发存活率和肿瘤学可行性。我们用阿替唑单抗和自体cevumeran治疗了16例患者,然后用mFOLFIRINOX治疗了15例患者。自体cevumeran在基准时间的3天内给药,是可耐受的,并在16名患者中的8名中诱导了新的高强度新抗原特异性T细胞,其中一半靶向一种以上的疫苗新抗原。使用一种新的数学策略来跟踪T细胞克隆(CloneTrack)和功能检测,我们发现疫苗扩增的T细胞占所有血液T细胞的10%,用疫苗加强剂重新扩增,并包括长寿命的多功能新抗原特异性效应物CD8+T细胞。在中位数为18个月的随访中,具有疫苗扩增T细胞的患者(应答者)比没有疫苗扩增T细胞的患者(无应答者)具有更长的中位无复发生存期(未达到);13.4个月,P= 0.003)。患者免疫适应能力的差异并没有混淆这种相关性,因为应答者和无应答者对同时发生的无关新型冠状病毒mRNA疫苗产生了同等的免疫力。因此,佐剂阿替唑单抗、自体cevumeran和mFOLFIRINOX诱导显著的T细胞活性,这可能与延迟的PDAC复发相关。
主要的
PDAC是美国第三大癌症死亡原因,世界第七大癌症死亡原因。随着发病率的增加,以及近60年来12%的存活率基本保持不变,预计到2025年,PDAC将导致更多的全球癌症死亡。手术是治疗PDAC的唯一方法。然而,尽管进行了手术,近90%的患者在7-9个月的中位时间内出现疾病复发,5年总生存率(OS)仅为8–10%。虽然辅助多剂化疗可延缓复发,并且是手术切除PDAC的标准治疗,但近80%的患者在14个月左右会复发,并且他们的5年OS < 30%。放射疗法、生物制剂和靶向疗法也无效。
PDACs对免疫检查点抑制剂几乎完全不敏感(< 5%反应率)。这种不敏感性部分归因于这样一个事实,即PDACs具有低突变率,产生很少的新抗原,这种突变产生的蛋白质在健康组织中不存在,将癌症标记为T细胞的外来物,因此潜在地使PDACs具有弱抗原性,很少有浸润的T细胞。然而,最近的观察表明,大多数PDACs实际上含有比以前预测的更多的新抗原。此外,对PDAC长期存活者的研究表明,新抗原可能刺激PDAC的T细胞。富含免疫原性新抗原的原发性肿瘤还具有大约12倍高密度的活化CD8+ T细胞,这与延迟的疾病复发和更长的患者生存期相关。因此,由于大多数PDACs含有具有刺激T细胞潜力的新抗原,递送新抗原的策略可能诱导新抗原特异性T细胞并影响患者结果。
基于观察到PDAC的长期存活者自发产生针对肿瘤特异性新抗原的T细胞反应,而这些新抗原在患者之间并不共享,我们测试了佐剂个体化疫苗是否能刺激新抗原特异性T细胞,并为手术切除PDAC的患者提供临床益处。治疗性mRNA疫苗技术促进了完全整合到常规肿瘤工作流程中的个体化新抗原疫苗的快速递送。此外,mRNA可以快速制造成具有多种新抗原的个体化疫苗,可以激活抗原呈递细胞并且可以使用新开发的临床阶段制剂有效地递送。因此,我们假设有效的个体化mRNA疫苗将在PDAC诱导新抗原特异性T细胞,消除微转移并延迟复发。
为了验证这一假设,我们进行了一项由研究者发起的I期临床试验,即序贯佐剂阿替唑单抗(基因泰克)、自体cevumeran(一种个体化的mRNA新抗原疫苗,在静脉内递送的lipoplex纳米颗粒中含有多达20种主要组织相容性复合体I类(MHCI)和MHCII类(MHCII)限制性新抗原;可手术切除PDAC患者的个体化新抗原特异性治疗(iNeST)、BioNTech和Genentech)和mFOLFIRINOX(图.1a)以:(1)扩增被PD-1信号传导抑制的新抗原特异性T细胞;和(2)使初次T细胞接触疫苗新抗原。
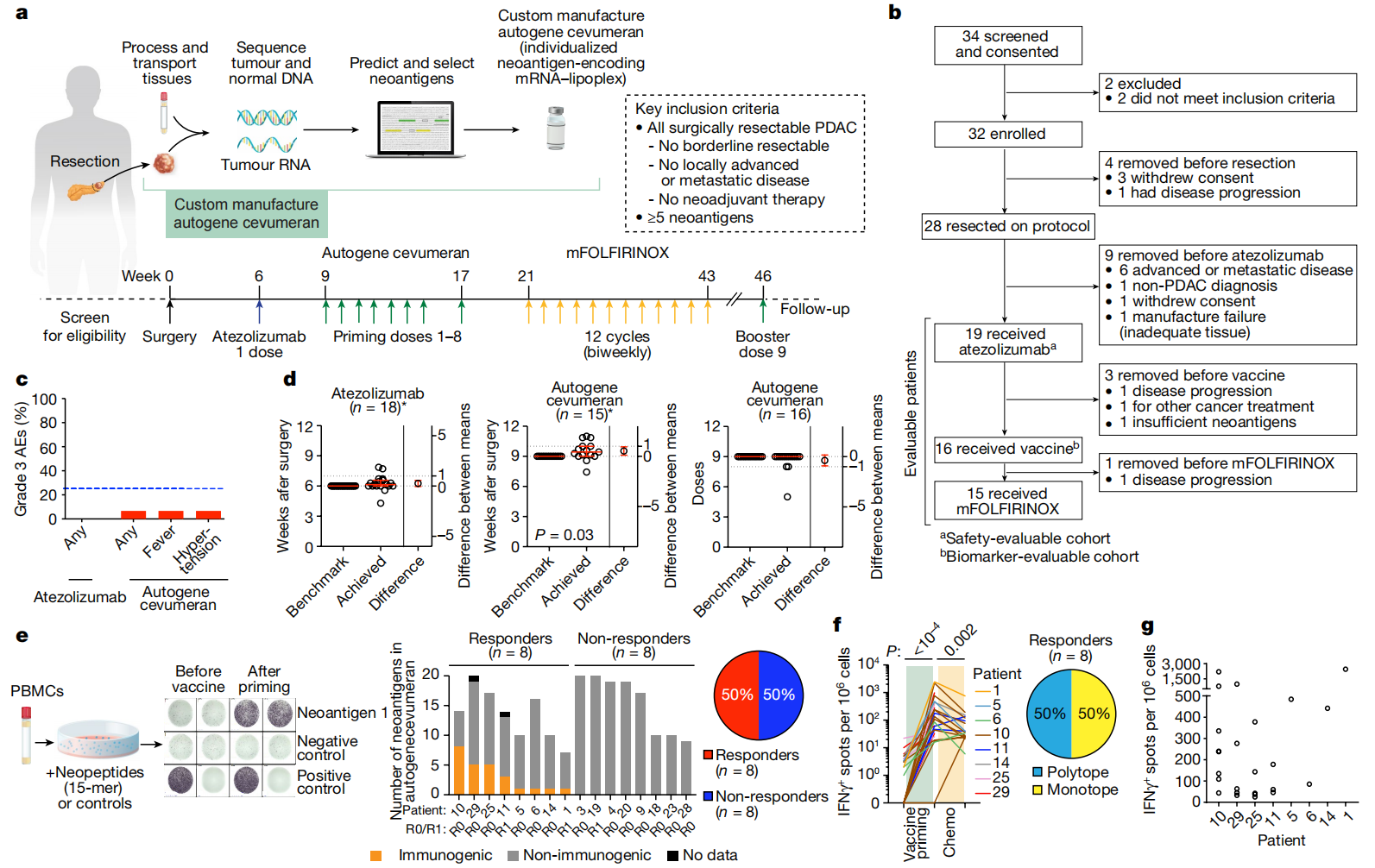
图1:个体化mRNA新抗原疫苗在PDAC患者中是安全、可行且具有免疫原性的。a,b、试验设计(a)和报告试验综合标准图(b). c,可归因于阿替唑单抗和自体cevumeran(疫苗)的3级AE在阿替唑单抗中的百分比(n= 19)和疫苗(n= 16)安全性可评估的患者。蓝线表示研究定义的安全阈值(25%)。d,达到和基准化的阿唑单抗时间(左)和第一次疫苗剂量(中),以及疫苗剂量数(右)。红线表示中值,误差线表示95%的置信区间,虚线表示临床无差别区。星号表示接受研究指定治疗序列的患者。e–g,对阿替唑单抗后和疫苗给药前、疫苗启动后1-3周和mFOLFIRINOX后5-6周收集的PBMCs进行IFNγ分析+通过体外IFNγ ELISpot检测所有单个疫苗新抗原的特异性T细胞n=生物标记可评估队列中的16名患者。如果ELISpot检测到IFNγ,患者被分类为应答者+针对至少一种疫苗新抗原的T细胞诱导。e,左,体外IFNγ ELISpot的示意图和代表性图像。中值,每位患者诱导IFNγ的疫苗新抗原数量+疫苗引发后收集的PBMCs中的T细胞。R0/R1表示手术边缘状态。对于患者25,针对2个新抗原库(库1具有2个新抗原,库2具有5个新抗原)检测到5个ELISpot反应中的2个。对,疫苗应答者和无应答者的比例。f,g,诱导新生反应的疫苗新抗原的标准化离体IFNγ ELISpot计数(n= 8名患者中的25个新抗原):纵向(f,左);启动后(g).减去未受刺激对照组的斑点计数。单表位与多表位患者对所有疫苗新抗原应答的比例(f,对)。n表示单个患者。化疗,化疗(mFOLFIRINOX)。P使用双尾不成对计算的值t-测试(d)或Wilcoxon配对符号秩检验(f).
安全性、可行性和免疫原性
从2019年12月到2021年8月,我们招募了34名患者,其中28名患者(图.1b)接受了手术。然后我们用阿替唑单抗治疗了19名患者,其中16名患者接受了自体cevumeran。这16名患者中有15名也接受了后续的mFOLFIRINOX治疗(图.1b).我们分析了安全性可评估队列的安全性(n= 19名接受阿替唑单抗治疗的患者,n= 16用自体基因cevumeran治疗),我们在一个生物标记可评估的队列中将免疫应答与RFS相关联(n= 16名接受阿替唑单抗和自体cevumeran治疗的患者)。所有19名可评估参与者都具有可切除PDAC患者的典型临床特征(扩展数据图.1a)。所有患者在纪念斯隆-凯特琳癌症中心(MSK)接受治疗和随访。
在安全性可评估队列中,接受阿替唑单抗治疗的19名患者中无1名出现3级或更高级别的不良事件(AE;图.1c)。在安全性可评估队列中,接受自体cevumeran治疗的16例患者中有1例(6%)出现3级AE(发热和高血压;图.1c)。接受自体cevumeran治疗的所有16名患者(100%)均出现1-2级AE(扩展数据图)。1c)。我们分别在基准时间的1天和3天内的中位时间给予阿替唑单抗和自体cevumeran(阿替唑单抗的中位时间为6.1周(范围为4.3-7.9周);自体cevumeran的中位时间为9.4周(范围为7.4-11.0周);图.1d)。19名患者中只有1名(5%)的新抗原不足,导致疫苗无法生产(图.1b)。16名患者中有3名(19%)没有接受全部9剂疫苗(图.1d),这是由于进展、死亡或mFOLFIRINOX毒性。因此,即使在复杂的肿瘤手术后,自体cevumeran也可以快速给药。
接下来,为了测量自体cevumeran诱导的T细胞反应,我们利用了先前描述的体外IFNγ ELISpot分析它可以检测对疫苗的高强度T细胞反应,而不区分CD8+来自CD4+T细胞反应。接受疫苗的16名患者中有8名(50%)产生了T细胞应答,通过体外IFNγ ELISpot检测,被认为是自体cevumeran应答者(图.1e)。通过测试新抗原疫苗中包含的每个特定靶位,我们检测到在所有患者中施用的230个新抗原中的25个(11%)诱导了足够高强度的T细胞反应,可通过体外IFNγ ELISpot检测(图.1e和扩展数据图.2a)。接受疫苗的所有患者中有一半出现了针对至少一种疫苗新抗原的新抗原特异性T细胞反应(中位数= 2,范围= 1-8;扩展数据图.2b)。此外,这些反应中有一半是多型的,靶向一种以上的疫苗新抗原(图.1e,f)。在接种前,通过体外IFNγ ELISpot检测不到针对疫苗新抗原的T细胞反应(图.1f)。疫苗接种后检测到的新抗原特异性免疫反应水平范围从每百万个大体积外周血单核细胞(PBMC)约100个斑点到每百万个大体积PBMC > 2000个斑点(图.1g)。观察到所有反应的数量和大小在患者间存在差异,多形反应的大小在患者内存在差异(图.1g).因此,自体cevumeran在大部分PDAC患者中诱导了大量的新生T细胞反应。
T细胞克隆型和表型
为了使用正交技术确定ELISpot试验的反应性,并探索自体扩增T细胞克隆的多样性和特异性,我们开发了CloneTrack。CloneTrack是一种新的数学和免疫学方法,它使用治疗前后外周血样本的T细胞受体(TCR) Vβ测序来识别治疗扩展的高量级T细胞克隆。使用克隆追踪,我们在8个应答者中的8个和8个无应答者中的1个中检测到疫苗诱导的克隆扩增(图.2a,b和扩展数据图.2d)。在应答者中,自体cevumeran扩增了多个克隆(平均7.5个克隆;图.2c)从检测不到的水平到高达10%(中位数为2.8%;图.2c)的所有血液T细胞。类似地,我们将CloneTrack应用于阿替唑单抗治疗前后收集的外周血样本,发现自体cevumeran扩增的T细胞克隆与阿替唑单抗扩增的T细胞克隆不重叠(扩展数据图.3a–c)。为了评估自体-cevumeran扩增的T细胞克隆的抗原特异性,我们在体外鉴定了对ELISpot鉴定的免疫显性新抗原特异的T细胞克隆,并在2名有单一应答的患者和2名有多型应答的患者体内检测了与自体-cevumeran扩增的克隆的克隆重叠(图.2d和扩展数据图.4a、b)。四分之三的患者(75%)和51%(n= 41个克隆中的21个)含有免疫显性新抗原特异性克隆(图.2e)。在第四个患者(患者11)中,免疫显性新抗原特异性克隆处于较低水平的疫苗扩增克隆池中(扩展数据图.4b).因此,自体cevumeran在PDAC从头扩增多克隆新抗原特异性T细胞。
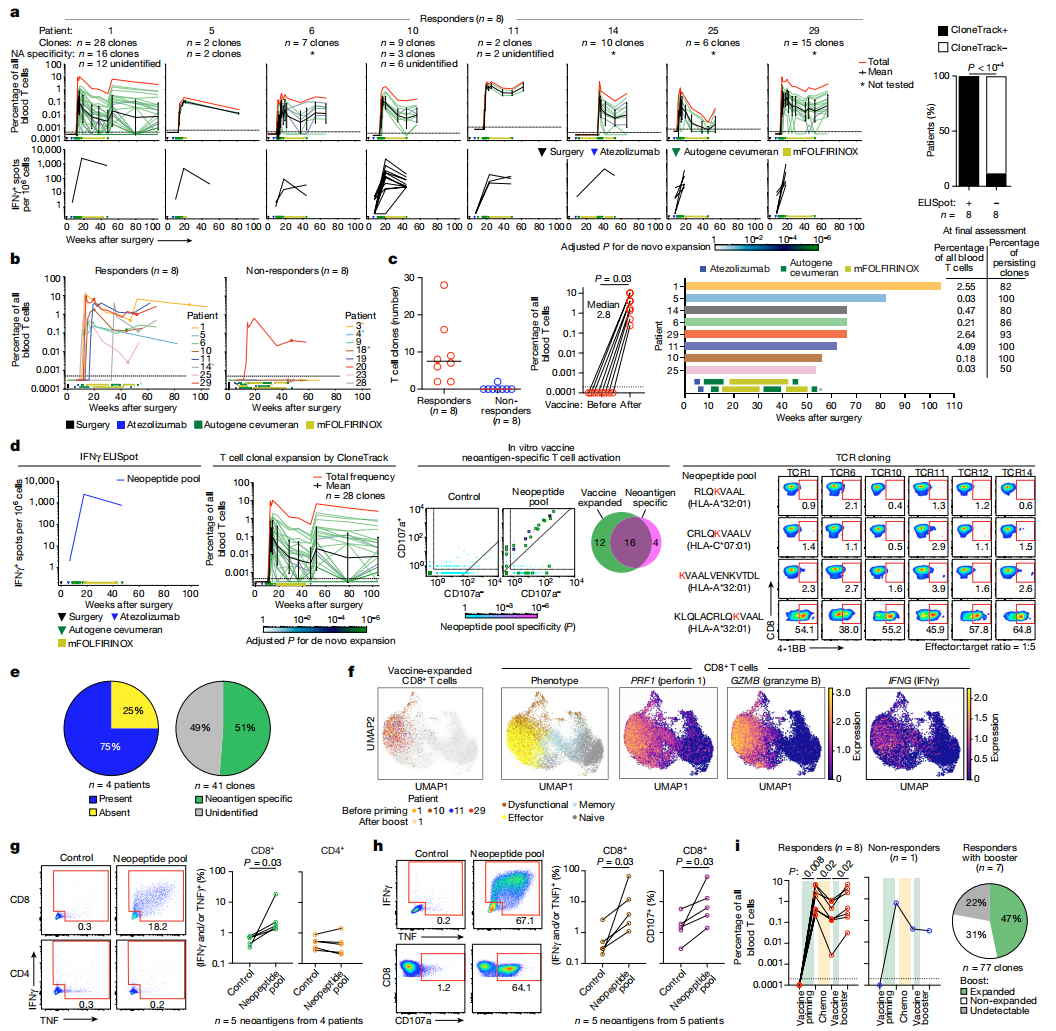
图2:扩增多克隆、多功能效应物CD8+T细胞的mRNA疫苗。a,使用CloneTrack评估的疫苗扩增的T细胞克隆(上),通过ELISpot评估的疫苗诱导的IFNγ(下)及其相关性(右)。b,c,通过克隆追踪鉴定的疫苗扩增克隆:纵向聚集百分比(b),唯一克隆的数量(c,左),疫苗和峰值扩张前聚集百分比(c、中间值)和每个患者的最终评估时间(条形图),以及最终评估时的聚集百分比和克隆分数(c,对)。为b,倒三角形表示单细胞测序的收集时间f圆圈表示疫苗加强剂。d免疫显性疫苗新抗原特异性克隆与疫苗扩增克隆重叠,以及通过患者1的TCR克隆对免疫显性疫苗新抗原的特异性。e,左,在疫苗扩增克隆中具有免疫显性疫苗新抗原特异性克隆的患者的百分比。正确,针对免疫显性疫苗新抗原的疫苗扩增克隆的百分比。f疫苗扩增的CD8的单细胞表型+t细胞。圆点表示血液中的CD8+t细胞。彩色点(最左边)表示疫苗扩增的克隆a. g,h,干扰素γ的百分比+,肿瘤坏死因子+和CD107a+CD8+ (g,h)和CD4+t细胞(g)在用离体免疫显性long(g)或最小(h)新肽再挑战。来自患者1的代表性血流图(g,h).CD3上孕+CD56–CD8+ (g,h)或CD4+ (g)细胞。i左,外周血中经过引发、化疗和加强的疫苗扩增克隆的聚集百分比。正确,用增强剂再次扩增的引物克隆的百分比。n表示单个克隆或患者。在…里a和d绿线表示单个克隆的轨迹;黑线,几何平均克隆轨迹(误差棒是几何标准差);红线表示所有扩展克隆的累积百分比。在…里b,c,星号表示改变的治疗计划。在…里b–d矩形表示治疗顺序。P使用修正的双尾费希尔精确检验计算的值(a,d,左),双面卡方检验(a,右),双尾配对t-测试(c),带Bonferroni校正的单尾二项式检验(d中)或双尾Wilcoxon配对符号秩检验(g–i).
我们接下来研究了自体cevumeran扩增的T细胞的表型和功能。使用单细胞RNA测序,我们发现自体cevumeran扩增的高量克隆是CD8+T细胞表达溶解标志(穿孔素1和颗粒酶B)和细胞因子(IFNγ),类似于保护性病毒疫苗诱导的效应T细胞(图.2f和扩展数据图.5a–c)。疫苗接种后收集的外周血样本始终含有多功能CD8+T细胞,但不是CD4+产生细胞因子(IFNγ和TNF)的T细胞在体外用两种长新肽再次攻击后脱颗粒(图.2g和扩展数据图.5d)和MHCI限制性最小表位(图.2h和扩展数据图.5e)。值得注意的是,尽管接种后mFOLFIRINOX治疗,自体cevumeran扩增的T细胞保持功能,并持续产生IFNγ(图.1f),在所有应答者中用疫苗加强剂进行统一的再扩张(图.2i)并在手术后2年内持续存在高达2.5%的所有血液T细胞(图.2c和补充表格1)。此外,疫苗加强剂在接受加强剂的7名患者中的7名中重新扩增了相同的致敏克隆(所有致敏克隆的47%;图.2i和扩展数据图.6)。尽管自体cevumeran扩增了多个克隆,但标准流式细胞术并不能可靠地检测T细胞的扩增和活化(扩展数据图)。5f)。总的来说,自体cevumeran大大扩展了T细胞,包括疫苗新抗原特异性、功能性和持久性CD8+T细胞。
疫苗反应和临床结果
在超过预先指定的次要终点的18个月的中位随访中,安全性可评估队列中的患者的中位OS和RFS未达到(图.3a)。对于生物标志物可评估队列中的患者,8名自体cevumeran反应者的中位RFS未达到,而8名无反应者的中位RFS为13.4个月(P= 0.003,危险比(HR) = 0.08 (95%可信区间(CI)0.01–0.4);图.3b)。为了排除响应时间偏差,我们进行了一项里程碑式的分析,将RFS与完成所有8次自体cevumeran初始剂量(里程碑式RFS)的无复发患者的反应相关联。与无反应者的11.0个月相比,有反应者同样没有达到中位界标RFS(P= 0.008,HR = 0.06(95% CI 0.008–0.40);图.3b)。与无反应者相比,反应者的血清CA19-9水平持续降低(扩展数据图.7a),最广泛使用的临床PDAC生物标志物。在生物标志物可评估的队列中,只有25%的患者在诊断时具有可检测的循环肿瘤DNA(扩展数据图.7b),如先前报道的可切除PDAC肿瘤患者,因此不是可靠的复发生物标志物。为了确定是否仅仅在预后较好的患者中增加了应答者,我们发现了对阿替唑单抗的应答、淋巴结阳性、切缘阳性、原发性肿瘤大小、化疗剂量数和瘤内CD8+T细胞密度与疫苗反应不相关(扩展数据图.1b和7c–e)。有反应者和无反应者也具有相当的免疫适应性,因为他们对与自体cevumeran同时施用的无关mRNA疫苗(新型冠状病毒)产生了等同的体液和细胞反应(扩展数据图)。8)。应答者和无应答者的所有主要先天免疫细胞和适应性免疫细胞的外周频率相同(扩展数据图.9a–c),以及相似的体细胞和种系遗传特征(补充表2–4)。总之,自体cevumeran扩增的T细胞反应与延迟的PDAC复发相关,这不会被患者选择、瘤内T细胞频率或外周T细胞频率或适应性的可检测差异所混淆。
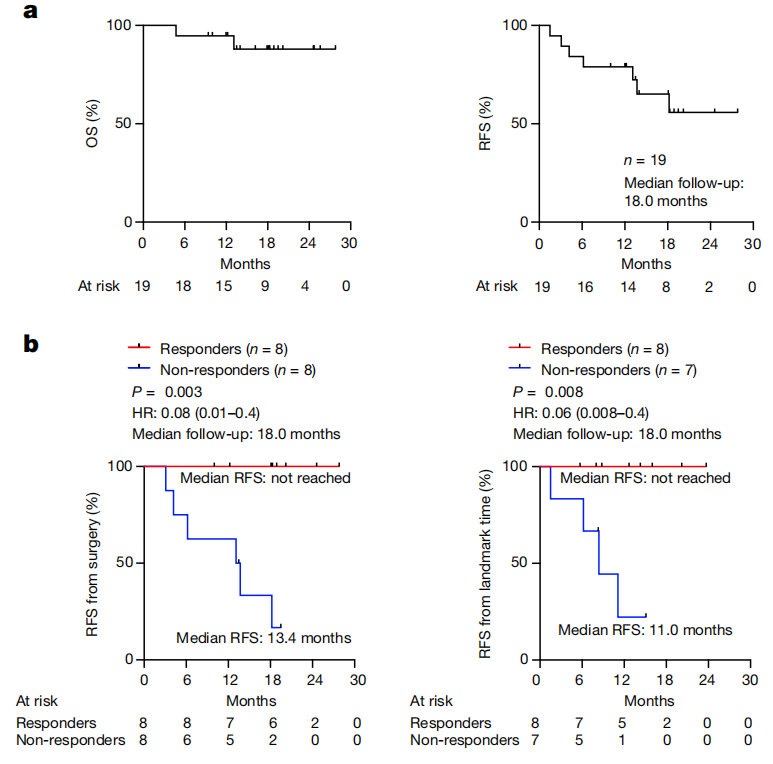
图3: mRNA疫苗反应与延迟的PDAC复发相关。a,OS和RFS在n=安全性可评估队列中的19名患者。b手术和里程碑时间(最后一次疫苗初始剂量的日期)的RFS,根据生物标记可评估队列中患者的疫苗反应进行分层。n表示单个患者。HR表示95%可信区间的危险比。P使用双尾对数秩检验计算的值。
由于自体cevumeran在应答者中诱导了对106个疫苗编码的新抗原中的25个(24%)特异的高水平T细胞应答(扩展数据图.2a),我们寻找疫苗反应的相关因素。我们之前的发现显示富含CD8+T细胞的PDAC肿瘤也富含免疫原性“高质量”新抗原,在肿瘤克隆中分布比例更大。因此,我们研究了肿瘤克隆性和新抗原质量是否与疫苗诱导的T细胞反应相关。一致地,自体cevumeran反应者比无反应者有更多的克隆性肿瘤(扩展数据图.10a)。接下来,我们检查应答者中的免疫原性新抗原是否包含高质量的特征。我们修改了之前描述的模型鉴定肿瘤中自发靶向的新抗原,以将免疫原性与接种的新抗原的质量相关联。在应答者中,新抗原质量作为一个连续变量与诱导IFNγ ELISpot反应的疫苗新抗原相关(扩展数据图.10b)。值得注意的是,无应答者与应答者具有相似数量的非同义突变和疫苗新抗原(扩展数据图)。2c)。
疫苗克隆和微转移
患者29对自体cevumeran有反应,其扩增的血液T细胞的最大百分比为第二高(图.2b)包括疫苗新抗原特异性多官能CD8+T细胞(扩展数据图.5d,e)。患者29的血清CA19-9水平升高,出现新的7mm肝脏损伤,提示疫苗启动后发生转移(图.4a)。活检样本没有显示恶性细胞,但密集的淋巴浸润(图.4b左)包括所有15个自体cevumeran-expanded(图.4b中)CD8+T细胞克隆具有裂解和效应潜能的表型证据的(图.4d)。数字液滴PCR显示,这种淋巴浸润含有罕见的细胞,这些细胞含有淋巴细胞。TP53R175H突变,与该患者原发肿瘤中的R175H驱动突变相同(图.4c和扩展数据图.10c)。肝脏病变在随后的成像中消失(图.4a),这表明自体扩增的T细胞可能具有根除微转移的能力。
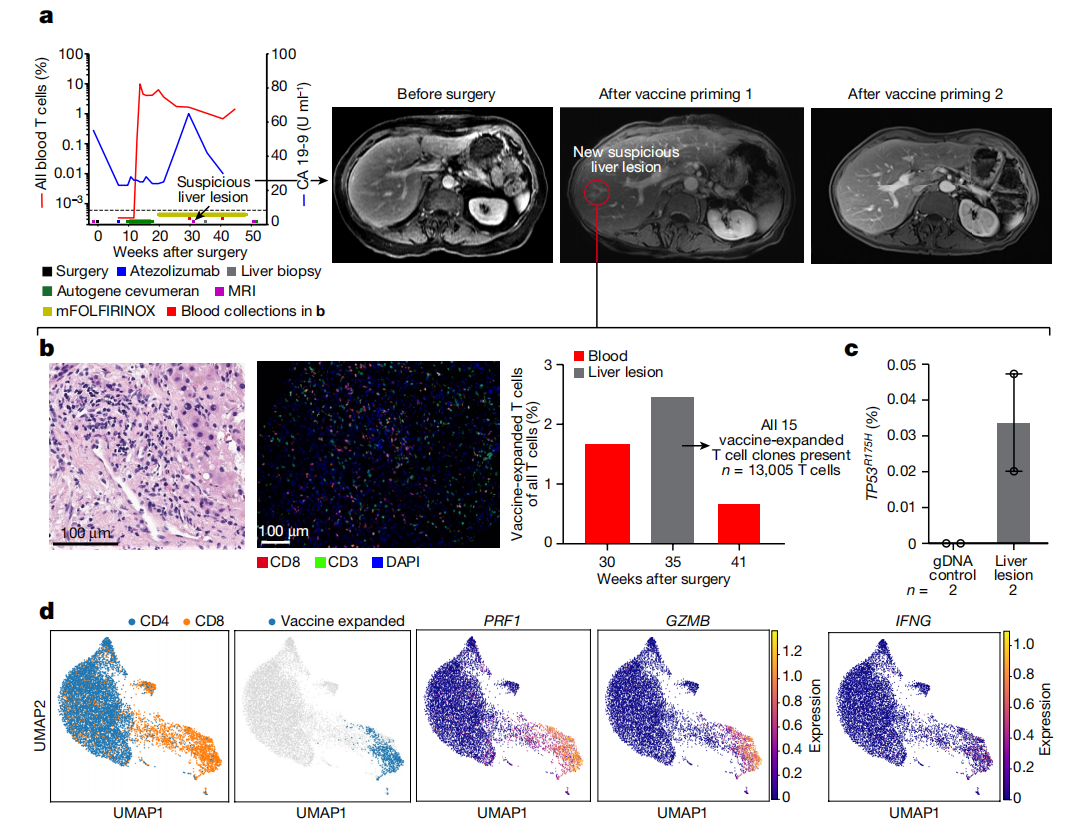
图4:疫苗扩增的T细胞可以渗入微转移灶。对疫苗有反应的患者接种疫苗后,肝内淋巴聚集消失的临床和免疫学快照。a使用CloneTrack和血清CA19-9(左)以及接种前后的腹部MRI(右)分析血液中疫苗扩增的T细胞的系列百分比。b、苏木精和曙红染色(左)、多重免疫荧光(中)和使用CloneTrack(右,灰条)测量的疫苗扩增的T细胞的百分比a。所有15个疫苗扩增的T细胞克隆(a红线)存在于肝脏损伤中(右侧,灰色条)。c,突变体的百分比TP53R175H数字化液滴PCR在肝脏病变中的应用。横条表示中值,误差横条表示标准差。d所有血液T细胞(左)和疫苗扩增克隆(中)的单细胞表型的一致流形近似和投影(UMAP)图,带有效应物标记(右)。n表示在肝脏病变中检测到的T细胞数量(b)或技术复制(c).数据代表对单个患者的分析。
讨论
我们证明了佐剂自体cevumeran(一种基于尿苷mRNA脂质体纳米颗粒的个体化新抗原疫苗)与阿替唑单抗和mFOLFIRINOX联合应用是安全可行的,并且在50%未选择的可切除PDAC患者中产生大量新抗原特异性T细胞。疫苗扩增的T细胞是持久的,尽管接种后mFOLFIRINOX治疗,仍可持续长达2年。高强度疫苗诱导的T细胞反应,是我们免疫反应分析的焦点,包括一种追踪疫苗扩增克隆的新方法,与延迟的PDAC复发相关。尽管样本量有限,这些早期结果保证了在PDAC对个体化mRNA新抗原疫苗进行更大规模的研究。
作为多重免疫疗法已经出现了免疫炎症性肿瘤,但是对于大多数患有非炎症性肿瘤的患者来说,仍然需要新的免疫疗法,这些患者对目前的免疫疗法基本上不敏感。事实上,普遍的想法是,这种肿瘤的低过客突变率使它们没有足够的新抗原用于疫苗。在这里,我们提供了证据,尽管PDAC突变率低,但mRNA疫苗可以诱导T细胞对这种癌症中新抗原的活性,这种癌症是一种主要具有免疫排斥或荒漠表型的非炎症性肿瘤。mRNA新抗原疫苗是否可以类似地激活其他非炎症性癌症中的T细胞,应该进行更广泛的测试。
我们没有发现疫苗应答与延迟复发的相关性被已知的预后变量(如淋巴结或边缘阳性疾病)混淆的证据。平均而言,无反应者的原发性肿瘤比有反应者略大;然而,较大的原发肿瘤大小与较短的RFS无关。由于尿苷mRNA脂复合物疫苗技术是基于有效的抗原递送到淋巴样区室,并刺激脾切除小鼠的弱T细胞反应值得注意的是,无应答者在脾切除患者中也略有增加(扩展数据图)。1b)。此外,疫苗在50%的患者中诱导了高水平的T细胞反应,这可能突出了对生物标志物的需要,以选择用于该治疗的最佳患者和肿瘤。值得注意的是,尽管自体cevumeran被设计成激活新抗原特异性CD4+和CD8+我们发现它激活了大量的CD8+T细胞在PDAC,本研究中的初级和确认性免疫应答试验不区分CD8+来自CD4+T细胞反应。事实上,由于这些分析偏向于高强度的T细胞反应,检测低强度反应的分析可能包括CD4+T细胞反应和预先存在的反应。在其他肿瘤中,我们观察到相当大比例的疫苗新抗原诱导的新生反应低于体外可检测的阈值,该反应水平未在本试验中评估。然而,我们在PDAC的研究结果表明,高水平的T细胞反应可能有助于获得良好的临床结果。因此,正在寻求确保高强度反应的策略,包括进一步优化mRNA疫苗效力和扩展新抗原发现空间,以包括除单核苷酸变异(SNVs)和插入和缺失(indels)之外的遗传异常(例如融合)。尽管如此,随着疫苗扩增多克隆T细胞,疫苗诱导的克隆多样性是否有助于持久控制是未来工作的另一个关键问题。
我们的研究不能检测疫苗反应的生物标志物的差异。尽管有这种限制,我们观察到应答者的肿瘤更具克隆性——可能代表免疫编辑进化中的肿瘤,如在长期存活者的免疫原性PDACs中所见。因此,我们推测,克隆性更强的原发性肿瘤可能反映了免疫系统识别肿瘤并因此对疫苗产生反应的能力。此外,观察到新抗原质量与免疫原性疫苗的相关性新抗原为选择的新抗原可能具有疫苗可能需要的更高免疫原性的概念提供了进一步的支持。然而,在本试验中,由于应答者和无应答者具有来自相似数量肿瘤突变的相似数量的疫苗新抗原,我们认为无应答者无应答不太可能是由于未能包括免疫原性新抗原。总的来说,这些观察仍然是初步的,但支持未来的研究,即肿瘤克隆性和新抗原质量是否可以作为疫苗反应的生物标志物。
我们在佐剂环境中测试了个体化mRNA癌症疫苗,这是由于观察到针对病原体的疫苗在历史上在预防而非治疗环境中最有效,这可能反映了疫苗效力需要最佳功能的宿主免疫系统。在晚期癌症患者中,宿主免疫的整体损伤和肿瘤之间新抗原异质性的知识差距可能会阻碍新抗原疫苗接种。因此,我们建议疫苗应在残留疾病最小的患者中进行测试,正如目前正在进行的高危结直肠癌试验(ClinicalTrials.gov标识符NCT04486378)和三阴性乳腺癌(ClinicalTrials.gov标识符NCT02316457)。值得注意的是,我们的研究表明,mRNA新抗原疫苗可以在9周内个体化,并完全整合到标准的临床工作流程中,即使在复杂的肿瘤手术后。鉴于该试验是PDAC个体化mRNA疫苗接种的重要经验,在手术后12周给予mFOLFIRINOX。此外,由于样本量有限,这项试验只招募了白人。未来的研究必须在PDAC病患者的不同人群中测试个体化的mRNA新抗原疫苗,以及更快的mFOLFIRINOX佐剂时间。个体化癌症疫苗的经验16先于并加速了基于mRNA的新型冠状病毒疫情疫苗38,现在可以进一步加快个体化癌症疫苗的生产时间,并实现更快速的辅助定制疫苗接种和化疗。。
总的来说,我们报告了初步证据表明,佐剂自体cevumeran(一种个体化的mRNA新抗原疫苗)与阿替唑单抗和mFOLFIRINOX联合使用,可在手术切除的PDAC患者中诱导显著的T细胞活性,这与延迟复发相关。一项后续全球随机试验(IMCODE 003,BNT122)即将进行。
Rojas LA, Sethna Z, Soares KC, Olcese C, Pang N, Patterson E, Lihm J, Ceglia N, Guasp P, Chu A, Yu R, Chandra AK, Waters T, Ruan J, Amisaki M, Zebboudj A, Odgerel Z, Payne G, Derhovanessian E, Müller F, Rhee I, Yadav M, Dobrin A, Sadelain M, Łuksza M, Cohen N, Tang L, Basturk O, Gönen M, Katz S, Do RK, Epstein AS, Momtaz P, Park W, Sugarman R, Varghese AM, Won E, Desai A, Wei AC, D'Angelica MI, Kingham TP, Mellman I, Merghoub T, Wolchok JD, Sahin U, Türeci Ö, Greenbaum BD, Jarnagin WR, Drebin J, O'Reilly EM, Balachandran VP. Personalized RNA neoantigen vaccines stimulate T cells in pancreatic cancer. Nature. 2023 May 10:1–7. doi: 10.1038/s41586-023-06063-y. Epub ahead of print. PMID: 37165196; PMCID: PMC10171177.
https://m.sciencenet.cn/blog-55647-1391655.html
上一篇:[转载]347.免疫印迹:了解新冠肺炎
下一篇:[转载]349.澳大利亚蝙蝠看护者狂犬病暴露前预防和加强剂量后的免疫反应