博文
《科学》:发现去乙酰化酶内源性阻断剂
||||
最近《科学》发表来自美国旧金山Gladstone研究所的一篇reports类型的论文,报道他们发现一种常见脂肪代谢产物β-羟基丁酸酯是去乙酰化酶的内源性阻断剂,这一发现对于解释β-羟基丁酸酯的一些奇怪效应提供了非常重要的证据,也给一些生物现象如运动、禁食、热量限制等代谢相关的生物学效应提供了更精确准确的解释。这一研究具有潜在的应用价值,例如或许可采用这种物质来代替热量限制、禁食和运动等苦行方式,来达到上述方法同样的健身和治疗疾病的效果。当然这仍需要更深入研究和针对这一物质开发出更理想合理的药物。
过去发现,细胞代谢产物如acetylCoA和还原型烟酰胺腺嘌呤二核苷酸(NAD)都是重要的基因表达辅助调节因子,影响表观遗传学修饰。例如他们可以调节组蛋白表达修饰乙酰转移酶(HATS)的活性依赖于核乙酰CoA浓度。Ⅲ类去乙酰化酶HDACs又被被称为沉默信息调节因子sirtuins,活性主要依赖于NAD的浓度。(从逻辑上考虑,一切代谢物都可能会变构调节相应酶活性,或者相关基因表达。)
HDACs共有四种类型,其中I类HDAC1,2,3,8,II类HDAC4,5,6,7,9,10和Ⅳ类HDAC11,这些酶都是锌依赖酶,但目前不清楚他们的内源性调节物是什么。(存在的问题,目前只知道Ⅲ类去乙酰化酶sirtuins的内源性调节物NAD)
I和II类HDACs的小分子抑制剂包括丁酸,丁酸细菌厌氧发酵的产物。β-羟基丁酸酯和丁酸的结构非常类似,β-羟基丁酸酯是高等动物中的脂肪酸氧化代谢产物酮体(图1A),在长时间运动或饥饿时哺乳动物将酮体作为如脑等许多器官的主要能量来源。禁食期间,肝脏把葡萄糖切换为脂肪酸氧化,血液中增加β-羟基丁酸酯1至2mM,长时间的禁食甚至更高(6-8 mM),在糖尿病酮症酸中毒可以达到> 25毫摩尔。(背景介绍)
为了测试β-羟基丁酸酯可能具有HDAC抑制剂活性,我们选择人类胚胎肾细胞(HEK293)用不同量的β-羟基丁酸酯处理,通过Western印迹测量乙酰化的组蛋白H3的赖氨酸9(AcH3K9)和组蛋白乙酰化水平。结果发现,β-羟基丁酸酯可以剂量依赖性提高组蛋白乙酰化水平。即使在1至2mM浓度(可以发生在人类后2-3天快速或剧烈运动后)仍具有显著效果,而且和丁酸的作用类似,不影响α-微管蛋白的乙酰化,说明能够抑制Ⅰ类HDACs,对 IIb类微管蛋白去乙酰化酶HDAC6没有作用。
为确定β-羟基丁酸酯的选择性HDAC抑制活性,在人类HEK293T细胞采用表位标记(FLAG)重组人HDAC1,HDAC3,HDAC4和HDAC6表达载体瞬时转染,纯化HDACs孵育[3H] - 标记的乙酰化组蛋白H4肽,检测脱乙酰基酶的活性。结果发现,β-羟基丁酸酯可以剂量依赖性抑制HDAC1,HDAC3,HDAC4,IC50分别为5.3,2.4,和4.5毫摩尔。但HDAC6 IC50高得多(48.5毫摩尔)。β-羟基丁酸酯对天然底物微管蛋白没有抑制HDAC6的作用。为测试组蛋白乙酰化水平提高的可能性,因为在靶组织中β-羟基丁酸酯可被分解为乙酰-CoA,直接测量在β-羟基丁酸酯处理的HEK293细胞,结果没有观察到显著变化。实验还测定了β-羟基丁酸酯是否对βOHBp300和PCAF(P300/CBP-相关因子)组蛋白乙酰转移酶活性,也没有发现任何变化。因此,毫摩尔浓度β-羟基丁酸酯直接通过HDAC抑制增加组蛋白乙酰化。在HEK293细胞中,高浓度的乙酰乙酸酯(乙酰丙酮)也可抑制体外I类和IIa类HDACs活性。但在禁食期间,乙酰乙酸酯在血液中只能达到β-羟基丁酸酯浓度的三分之一或更少。因此在生理条件下,乙酰乙酸酯不太可能达到抑制HDACs的浓度。β-羟基丁酸酯脱氢酶1和2能使β-羟基丁酸酯转化为乙酰乙酸,为测试β-羟基丁酸酯、乙酰乙酸和乙酰-CoA在β-羟基丁酸酯组蛋白乙酰化阻断效应中的相对贡献,用siRNA阻断β-羟基丁酸酯脱氢酶1和2的表达,结果发现对3mM的β-羟基丁酸酯的组蛋白乙酰化作用没有影响。但可抑制较高的浓度10和30毫摩尔β-羟基丁酸酯,说明是乙酰乙酸和乙酰-CoA对IC50以上浓度的效应中有一定贡献。(这个部分说明β-羟基丁酸酯对组蛋白乙酰化的影响有一定选择性)
为确定β-羟基丁酸酯体内的浓度变化可能会影响组蛋白乙酰化,研究测量小鼠血清中的β-羟基丁酸酯浓度后,对动物进行24小时禁食或热量限制饮食,另外采用渗透泵给动物腹腔注射β-羟基丁酸酯或PBS对照。结果发现,禁食24小时β-羟基丁酸酯浓度增加至1.5±0.1Mm,热量限制动物0.6±0.1 mM,注射β-羟基丁酸酯动物1.2±0.1 mM。
H3K9和H3K14乙酰化反映了人类在内多种物种HATs和HDACs竞争活性,采用免疫杂交检测组织组蛋白乙酰化水平,结果发现禁食小鼠肾等器官组蛋白乙酰化显著增加,在热量限制小鼠肾组蛋白乙酰化增加2〜5倍。肾组蛋白乙酰化血清β-羟基丁酸酯水平呈现正相关关系,其中和组蛋白H3K9相关系数R 2 =0.772,H3K14相关系数R 2= 0.863。随后针对组蛋白乙酰化最大变化的肾组织,作为β-羟基丁酸酯影响基因表达和细胞表型的器官。
将HDAC抑制剂诱导的组蛋白乙酰化与基因转录激活和抑制属于一个家族。为分析β-羟基丁酸酯对该系统的影响,从β-羟基丁酸酯治疗和PBS对照处理后24小时小鼠肾脏组织提取mRNA,进行微阵列分析。在35,556个目标基因中,284个表达上调,其中表达上调最显著的5个基因中,4个基因在禁食的微阵列分析和PCR结果同样上调。
采用生物通路和网络分析工具Ingenuity Pathway Analysis (IPA)。
确定了两个β-羟基丁酸酯的诱导基因MT2和LCN2(受到FOXO3A调控)。通过定量PCR,发现属于FOXO3A网络的5个基因(FOXO3A,MT2,LCN2,Lemd3和HBP1)均受到β-羟基丁酸酯的诱导。
FOXO3A是一种转录因子,诱导细胞周期停滞和抗氧化应激反应。通过定量PCR发现,金属硫蛋白2(MT2)是β-羟基丁酸酯诱导表达最明显的基因,是减少氧化应激的重要分子。热量限制可以小幅增加FOXO3A和MT2 mRNA。染色质免疫沉淀法(ChIP)分析发现,β-羟基丁酸酯10毫摩尔处理可以使HEK293细胞FOXO3A和MT2启动子H3K9乙酰化水平提高。
接下来,选择性shRNA阻断每个I类和II HDAC表达,每个HDAC的shRNA的选择性抑制60%同源HDAC的表达。结果发现, HDAC1或HDAC2的表达抑制后, FOXO3A mRNA上调 1.8倍,MT2 mRNA 上调1.5倍。如果HDAC1或HDAC2的表达同时被抑制,FoxO3a增加和MT2的mRNA上调更明显。ChIP分析显示,HDAC1(而非HDAC6)可以结合到FOXO3A和MT2启动子上。β-羟基丁酸酯处理不影响HDAC1和启动子的结合,提示β-羟基丁酸酯是促进局部氧化应激抗性基因FOXO3A和MT2的启动子的组蛋白乙酰化水平。
线粒体超氧化物歧化酶(Mn-SOD)和过氧化氢酶(CAT)也是FOXO3A目标基因,也属于氧化应激保护蛋白。研究结果发现β-羟基丁酸酯处理可以使小鼠肾组织Mn-SOD和CAT表达增加。
MT2、FOXO3A、Mn-SOD和CAT表达增加可能是β-羟基丁酸酯抗氧化应激的内在原因。羰基衍生物是金属催化氧化蛋白质氨基酸残基如脯氨酸、精氨酸、赖氨酸和苏氨酸的产物。羰基化是不可逆、不可修复并具有积累效应。为了证明β-羟基丁酸酯的抗氧化作用,先给动物腹腔连续注射24小时,静脉注射百草枯2小时后,处死动物去肾脏组织进行DNP蛋白免疫分析以确定羰基化水平,结果发现β-羟基丁酸酯可以显著降低百草枯引起的肾脏组织蛋白羰基化。
根据上述研究结果,可以判断β-羟基丁酸酯是一种内源性HDAC抑制剂,当动物长时间禁食和热量限制时,体内β-羟基丁酸酯的增加(毫摩尔级别)可以影响基因表达的改变,这是一种代谢和基因相互关联的新模式。研究发现,β-羟基丁酸酯可保持组蛋白乙酰化状态,影响基因表达,主要发生在肾脏,或者说对肾脏的抗氧化具有更强的修饰作用(提示)。将来的研究应探讨β-羟基丁酸酯影响的更特异性基因表达和对其他器官组织的生理影响。例如,低碳水化合物的饮食,诱导生酮具有广泛的神经保护作用,提高神经元抗氧化损伤能力。此外,由基因操作或化学抑制HDAC活性可促进果蝇寿命延长。β-羟基丁酸酯抑制HDACs可能是生酮饮食的有益作用的机制之一,而限制热量摄入对健康的好处也得宜于这一途径。
博主点评,这一研究能发表在《科学》上,到底有什么重大创新?β-羟基丁酸酯的作用早就有人报道,本研究唯一发现的新内容是β-羟基丁酸酯具有选择性阻断HDACs的作用。如果按照作者提供的说法,他们也是根据细菌代谢产物丁酸具有这样的作用联想到的,那么似乎也没有什么高深的理论创新。研究的手段也都属于常规方法。从简易程度上考虑,作者或许是首先用β-羟基丁酸酯注射,观察对氧化损伤模型百草枯诱导的蛋白氧化损伤是否有影响,结果发现很有效果,然后根据β-羟基丁酸酯和丁酸结构的类似性,寻找β-羟基丁酸酯抗氧化是否是因为影响到HDACs,采用细胞学方法,证明确实具有类似作用,而且具有比较好的选择性。表达芯片应该是在后期做的,主要是希望寻找到HDACs的调节基因。其实这些似乎复杂的检测都是可以联想到的,几乎是必须应该检测的内容。《科学》大概就是这样的水平,每周那么多文章,不可能篇篇特别高的质量。
延伸阅读:组蛋白的可逆共价修饰调节是基因表达调控的重要方式之一。目前已知的共价修饰调节作用有六种修饰方式:磷酸化/ 去磷酸化、乙酰化/ 去乙酰化、腺苷酰化/ 去腺苷酰化、尿苷酰化/ 去尿苷酰化、甲基化/ 去甲基化、氧化(S-S)/ 还原 (2SH)。而首次证实组蛋白乙酰化/去乙酰化与转录活性密切相关的是十几年前发现的常染色质进行活跃转录的核心组蛋白是高度乙酰化的,而进行不活跃转录的核心组蛋白是低乙酰化的,经进一步研究表明:细胞通过对核心组蛋白进行可逆共价修饰来调节其N端尾部的乙酰化水平来调控转录的起始。随着对组蛋白在基因表达调控中所起的可逆共价修饰的深入研究,人们认识到乙酰化/去乙酰化是众多共价修饰中最重要的一种。
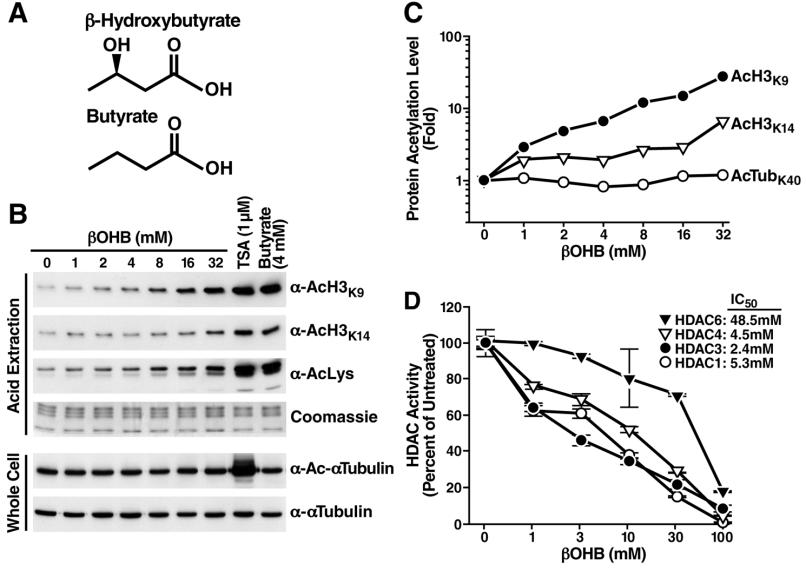
图1 在体和离体阻断效应
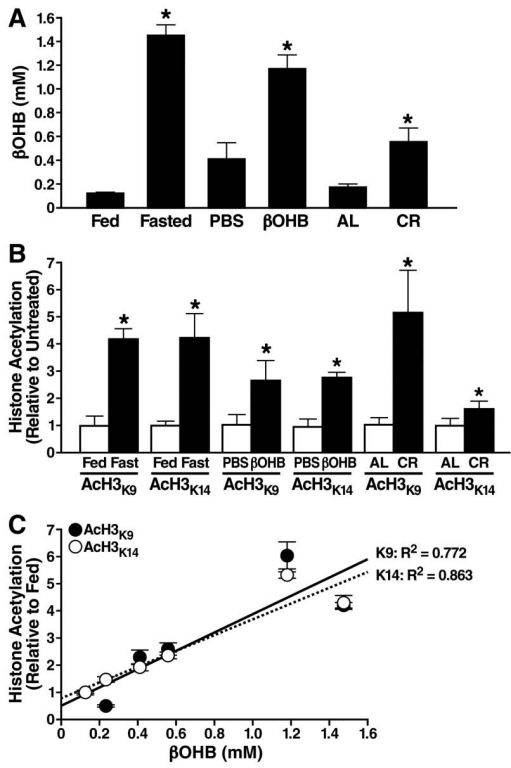
图2 禁食对血中β-羟基丁酸酯水平肾脏乙酰化的影响
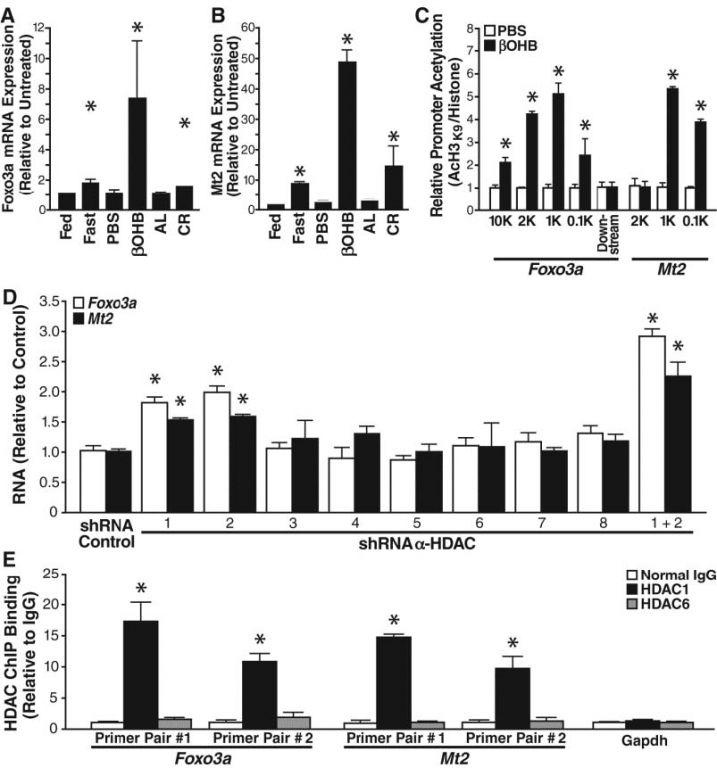
图3 β-羟基丁酸酯对细胞抗氧化能力的影响

图4 β-羟基丁酸酯对抗氧化的促进作用
https://m.sciencenet.cn/blog-41174-645326.html
上一篇:β羟丁酸是长生不老药
下一篇:中国学者在CNS杂志发表论文统计