博文
音乐起源之谜(四) ——音乐起源的性选择假说及其不足
 精选
精选
||||
音乐起源之谜(四)
——音乐起源的性选择假说及其不足
前言:上一节介绍了音乐的遗传基础,如果音乐是自然选择而来,那么促使音乐进化的“选择压”是什么?换句话说,音乐能给人带来什么好处?目前已有的一些假说认为,音乐可以吸引异性、促进母婴联系、增加社会凝聚力。虽然这些假说都各有道理,但是也各有不足,本节将逐一介绍。
前几节介绍了音乐现象的概貌。从行为学水平来看,音乐不仅广泛存在于人类历史和文化 (d'Errico et al., 2003; Conard et al., 2009),甚至某些动物也具有类似人类的音乐才能 (Patel et al., 2009; Schachner et al., 2009; Cook etal., 2013);从认知和神经生物学水平来看,音乐不但具有触发人类奖赏系统及诱导情绪的作用 (Blood et al., 1999; Blood and Zatorre, 2001; Menon and Levitin, 2005; Koelsch et al., 2006; Wieczorkowska et al., 2006; Koelsch, 2014),还能影响小鼠的心理和生理 (Chikahisa et al., 2006; Angeluccia et al., 2007; Li et al., 2010; Uchiyama et al., 2012);从基因水平来看,音乐才能具有遗传基础 (Drayna, 2001; Peretz et al., 2003; Peretz et al.,2007; Tan et al., 2014)。音乐的上述特点符合适应性状的所有特征。
如果说音乐是由自然选择进化而来,那么需要回答的一个关键问题:音乐起源和进化的选择压是什么?达尔文认为,人类音乐和舞蹈是性选择的结果;也有科学家认为音乐起源的驱动力在于音乐促进了母婴联系;还有科学家提出音乐的作用在于增加社会凝聚力。音乐起源的“性选择假说”、“母婴联系假说”和“社会凝聚力假说”虽然各有合理的一面,不过都或多或少的存在一些不足之处。下面以“性选择假说”为重点,逐一介绍。
性选择(sexual selection)
《艺文类聚》里有个“东食西宿”的故事,齐人有个女儿,同时被东西两家的儿子追求,东家的儿子人丑家富,西家的儿子人帅家贫。父母决断不下,就让女儿拿主意,怕女儿害羞不好开口,就让她伸手示意,结果女儿两只手都伸了出来。父母就奇怪了,问她什么意思,女儿说:“东家吃饭,西家睡觉”。
这个故事常常被用来形容贪得无厌的人,不过想想老祖宗说的“食色,性也”也就觉得无可厚非了。关于择偶,还有很多标准比齐人之女更甚,小说里的“潘驴邓小闲”就不提了,现实中的“白富美”、“高富帅”比齐人之女还多一个标准,“有车有房,没爹没娘”更狠。在此我对上述故事和标准不作道德评价,我只想引出生物学上的“性选择”这个概念,生物学上的“性选择”是包括人类在内的动物为了繁衍更多更好的后代而进行的特殊“自然选择”。动物的“求偶”和人类的“择偶”,都有同性之间的互相竞争,和异性之间的互相选择,最终目的都是为了获得更多更好的交配权,从而繁衍出更多更好的后代。
性选择由达尔文提出,并且在《人类的由来和性选择》(The Descent of Man and Selection in Relation to Sex)(Darwin, 1871)一书中作了比较详细的阐述。动物之间为了争夺交配权而进行的斗争就是性选择,这种性选择一般在雄性动物表现明显。绝大多数雄性动物生产精子的成本低、数量大,而且很多雄性在生殖和养育中不需要付出,或者只付出极少的能量;所以雄性获得的交配机会越多,后代的数量就会越大。相反,雌性动物一生中排卵的数量有限,而且在生殖和养育的过程中,胎盘和乳汁会支出很高的能量;为了保证有限后代的优良品质,雌性必须对雄性进行选择。此外,雌性动物一般有固定的发情期,而雄性则随时待命。基于上述原因,很多物种的雄性动物会对交配权展开激烈的竞争,主要通过“同性竞争”和“异性吸引”两种方式进行。

图1. 通过“同性竞争”进行的性选择。A. 雄鹿之间为争夺交配权展开的斗争;B. 公羊之间为争夺交配权展开的斗争;C. 具有强劲鹿角和羊角的个体在斗争中获胜,获得交配权,将基因遗传给后代,我们今天看天的鹿角和羊角就是性选择的结果。图片来源:网络。
“同性竞争”是指以武力斗争和驱赶同性,某些有蹄类动物,比如雄鹿(图1,A)、公羊(图1,B)等强劲的鹿角和羊角就是性选择的结果。在交配季节来临之际,雄性个体之间轰轰烈烈地上演着一场又一场的“比武招亲”,强劲有力的角是制胜之本。胜利者通过斗争赶走了竞争者,让雌性没有选择的余地,从而顺理承章地获得更多的交配权(图1,C)。胜利者的基因遗传给后代,其雄性后代也拥有强劲的角,经过长期的自然选择,鹿角和羊角进化成我们今天看到的样子。

图2. 通过“异性吸引”进行的性选择。A. 雄性天堂鸟具有漂亮的羽毛和优美的舞蹈;B. 两只雄性天堂鸟为了获得雌性的青睐,正在竭力展示自己的羽毛和舞蹈; C. 雌性选择了最赏心悦目的一只作为交配对象,主管雄性天堂鸟漂亮毛色和优美舞姿的基因得到遗传。图片来源:网络。
如果说“同性竞争”的性选择是一场武斗,那么“异性吸引”的性选择可以算得上文斗了。这种“文斗”可以是比华丽的外表,比如雄孔雀的羽毛;也可以是婉转的歌声,比如很多鸣禽的叫声;也可以是优美的舞蹈,比如丹顶鹤的求偶舞。不过更多的时候是集中了好几样才能,比如天堂鸟(图2)。在求偶的竞争中,雄性天堂鸟会充分展示出华丽的外表和优美的舞姿,雌性可以从中选择令自己最赏心悦目的一只进行交配。于是,主管漂亮羽毛和优美舞姿的基因在雄性天堂鸟代代相传。
音乐起源的性选择假说(sexual selection hypothesis)
不管是以武力驱赶同性,还是以外表吸引异性,优胜者可以获得更多的交配权,从而繁衍出更多的后代。达尔文认为人类的音乐和舞蹈跟鸟类的鸣叫和舞蹈一样,是经过性选择而来的适应现象。换言之,音乐起源的性选择假说认为,音乐可以帮助我们吸引异性,从而在求偶中获得优胜(图3)。虽然现实生活中能歌善舞的男女更能得到异性的青睐,而且性选择学说也得到一些科学家的认同和发展 (Miller, 2000),不过更多科学家对音乐起源的性选择学术持保留、甚至反对意见 (Hagen and Bryant, 2003)。

图3. 音乐起源的“性选择假说”。达尔文认为,人类的音乐和舞蹈类似鸟类的鸣叫和舞蹈一样,是性选择的结果。A. 原始部落的歌舞活动;B. 西方社会的歌舞活动;C. 能歌善舞的个体获得交配权,音乐和舞蹈的基因得到遗传和进化。图片来源:网络。
音乐不仅可以吸引异性,而且能够吸引同性,迈克尔·杰克逊和布兰妮·斯皮尔斯的歌迷遍布全球,男女通吃。音乐不仅能吸引性成熟的育龄男女,而且能吸引青春期之前的儿童,甚至能影响胎儿。音乐不仅用于舞会、婚礼等和性吸引相关的场合,还经常用于战争、政治活动和宗教(包括很多要求教徒禁欲的宗教)。音乐这些不分男女老少和社会场合的特征,都是性选择假说所不能很好解释的 (Hagen and Bryant, 2003)。此外,Daniel等人通过问卷调查发现,音质作为问卷设计的15种择偶因素之一,被男性排在倒数第二位,被女性排在倒数第一 (Daniel et al., 1985)。由此可见音乐起源的性选择假说存在诸多问题。
音乐起源的母婴联系假说(mother-baby interaction hypothesis)
婴儿的生命历程开始于母亲的摇篮曲,就算没有摇篮曲,也会有母亲的呢喃。Dissanayake提出音乐起源的“母婴联系假说”,认为音乐起源于母婴之间的互动,音乐建立了母婴之间的情感联系,和性选择无关 (Dissanayake, 2000; Trehub, 2003)。Dissanayake认为在婴儿6月龄之前,母婴之间通过一系列的行为、语音、面部表情和动作进行互动,所以音乐并不是单独起源的,还伴随了“时间上的模式运动(temporally patterned movements)” (Dissanayake, 2000)。而且音乐的功能不仅在于吸引和愉悦个体,还在于协调参与者的情绪,于是促进彼此的联系。

图4. 音乐起源的“母婴联系假说”。Dissanayake认为音乐起源于灵长类和人类母婴之间的互动,音乐建立了母婴之间的情感联系,和性选择无关 (Dissanayake, 2000)。A. 黑猩猩妈妈在抚慰其幼子 (BBC, 2014);B. 绘画作品中的母亲和孩子(图片来源:网络)。
既往研究表明,身体运动在音乐节奏感知中扮演重要作用 (Phillips-Silver and Trainor, 2005),而且音乐和运动似乎还共享情绪表达系统 (Sievers et al., 2013)。母婴联系假说认为母婴之间的语音和动作是音乐起源的原因,该思路可以解释上述研究发现。不过母婴联系假说的对象仅限于灵长类和人类,并不能扩展到其它更多的物种,鹦鹉 (Patel et al., 2009)和海狮 (Cook et al., 2013)对音乐的感觉运动同步化(sensorimotor synchronization)并不能通过母婴联系来解释。另外,从基因水平来看,如果将来在其它物种找到与人类音乐才能相关基因同源的基因,将会是对母婴联系假说的巨大挑战。所以,音乐起源的母婴联系假说似乎比性选择假说的处境更加堪忧。
音乐起源的社会凝聚力假说(social cohesion hypothesis)
1984年Roederer提出了音乐起源的“社会凝聚力假说” (Roederer, 1984)。Roederer认为,音乐和舞蹈活动给人带来的好处是促进了人与人之间的交流,从而增加了社会凝聚力。群体活动,比如集体狩猎(图5,A)对个体和群体来说是互为辅助的,个体为群体出力,群体也为个体提供回报。经常的载歌载舞(图5,B)也能增加群体的凝聚力。
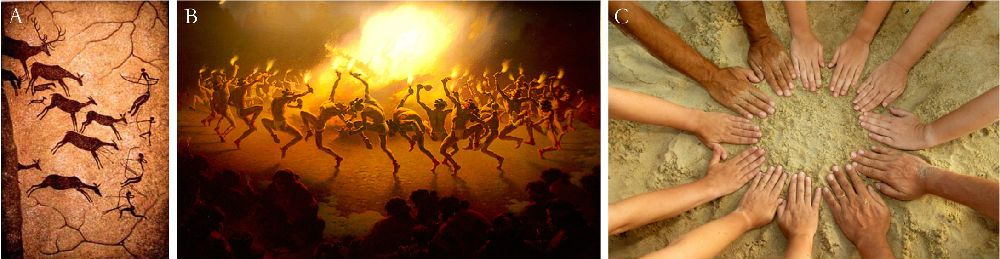
图5. 音乐起源的“社会凝聚力假说”(图片来源:网络)。Roederer认为音乐和舞蹈促进了人与人之间的交流,增加了社会凝聚力。A. 集体狩猎图;B. 原始的歌舞;C. 社会凝聚力。
该假说同样得到一些科学家的认可 (Brown, 2000; Freeman, 2000; Mithen, 2007),而且可以解释音乐和舞蹈在群体活动中的作用,不过仍然存在很大的争议 (Hagen and Bryant, 2003)。音乐能触发大脑的奖赏系统,社会凝聚力的回报似乎还没有强大到这种程度。同样的,音乐起源的社会凝聚力假说也面临着母婴联系假说一样的问题,那就是不能解释音乐对其它物种生理和行为的影响。
音乐起源需要更完善的假说
进化音乐学(evolutionary musicology)需要回答的关键问题是音乐进化的选择压问题,也就是音乐给人带来什么好处,使得与音乐才能相关的基因得到选择。虽然现有的音乐起源的“性选择假说”、“母婴联系假说”和“社会凝聚力”假说都能或多或少地解释一些音乐现象,不过仍然有很大的缺陷。关于音乐的起源,需要一个更加完善的综合性假说,这个假说需要对几乎所有的音乐现象给出全面而且深入的解释,而且这个假说给出的选择压需要强大到足够进化出音乐快感。最近我提出了音乐起源的“节奏适应假说”,从生物学和社会学角度,对音乐、舞蹈和语言的起源及进化勾勒出一条可能的路径,并为音乐的自然选择学说提出一个强有力的选择压——对内外环境中节奏事件的适应。该假说有望揭开音乐起源之谜,接下来的博文将详细介绍“节奏适应假说”,敬请关注。
注:1.该博文中凡有确切来源图片均已注明出处,其它图片收集自网络,原作者可以联系我撤销图片或添加备注。
2.成文仓促,难免失误,请大家多多指正;另,有交流或合作意向的老师和同学,欢迎随时联系。
王天燕(tianyanwang@hotmail.com)
2015年04月12日
相关链接:
A hypothesis on the biological origins and social evolution of music and dance (Wang, Front. Neurosci. 2015)
音乐起源之谜(一)——音乐、舞蹈和语言起源的“节奏适应假说” (王天燕,03/19/2015)
音乐起源之谜(二)——音乐的“文化起源” VS“生物起源” (王天燕,03/26/2015)
音乐起源之谜(三)——音乐才能的遗传基础 (王天燕,04/02/2015)
音乐起源之谜(四)——音乐起源的性选择假说及其不足(王天燕,04/12/2015)
音乐起源之谜(五)——音乐之始,大道至简 (王天燕,04/16/2015)
音乐起源之谜(六)——音乐和舞蹈统一于节奏运动 (王天燕,05/08/2015)
音乐起源之谜(七)——节奏适应是食物和性的前提 (王天燕,05/15/2015)
音乐起源之谜(八)——音乐在动物界的起源和进化 (王天燕,05/22/2015)
音乐起源之谜(九)——人类音乐的起源和进化 (王天燕,06/08/2015)
音乐起源之谜(十/完)——节奏适应假说的机遇和挑战 (王天燕,06/18/2015)
参考文献:
Angeluccia,F., Riccib, E., Paduaa, L., Sabinob, A., and Tonali, P.A. (2007). Musicexposure differentially alters the levels of brain-derived neurotrophic factorand nerve growth factor in the mouse hypothalamus. Neurosci. Lett. 429,152-155. doi: 10.1016/j.neulet.2007.10.005.
Bbc (2014). Did early humans, or even animals, invent music?doi: http://www.bbc.com/earth/story/20140907-does-music-pre-date-modern-man.
Blood, A.J., and Zatorre, R.J. (2001). Intensely pleasurableresponses to music correlate with activity in brain regions implicated inreward and emotion. Proc. Natl. Acad.Sci. U. S. A. 98, 11818-11823.doi: 10.1073/pnas.191355898.
Blood, A.J., Zatorre, R.J., Bermudez, P., and Evans, A.C.(1999). Emotional responses to pleasant and unpleasant music correlate withactivity in paralimbic brain regions. Nat.Neurosci. 2, 382-387. doi:10.1038/7299.
Brown, S. (2000). "Evolutionary models of music: fromsexual selection to group selection," in Perspectives in Ethology: Evolution, Culture, and Behavior, eds. F.Tonneau & N.S. Thompson. (New York, NY: Springer), 231-281.
Chikahisa, S., Sei, H., Morishima, M., Sano, A., Kitaoka, K.,Nakaya, Y., and Morita, Y. (2006). Exposure to music in the perinatal periodenhances learning performance and alters BDNF/TrkB signaling in mice as adults.Behav. Brain Res. 169, 312-319. doi:10.1016/j.bbr.2006.01.021.
Conard, N.J., Malina, M., and Münzel, S.C. (2009). New flutesdocument the earliest musical tradition in southwestern Germany. Nature 460, 737-740. doi: 10.1038/nature08169.
Cook, P., Rouse, A., Wilson, M., and Reichmuth, C. (2013). ACalifornia sea lion (Zalophus californianus) can keep the beat: motorentrainment to rhythmic auditory stimuli in a non vocal mimic. J. Comp. Psychol. 127, 412-427. doi: 10.1037/a0032345.
D'errico, F., Henshilwood, C., Lawson, G., Vanhaeren, M.,Tillier, A.M., Soressi, M., Bresson, F., Maureille, B., Nowell, A., Lakarra,J., Backwell, L., and Julienlo, M. (2003). Archaeological evidence for theemergence of language, symbolism, and music - An alternative multidisciplinaryperspective. Journal of World Prehistory17, 1-70. doi:10.1023/a:1023980201043.
Daniel, H.J., O'brien, K.F., Mccabe, R.B., and Quinter, V.E.(1985). Values in mate selection: A 1984 campus survey. College Student Journal.
Darwin, C. (1871). TheDescent of Man and Selection in Relation to Sex. London: John Murray.
Dissanayake, E. (2000). "Antecedents of the temporalarts in early mother-infant interaction," in The Origins of Music, eds. N.L. Wallin, B. Merker & S. Brown. (Cambridge, MA: MIT Press), 389-410.
Drayna, D. (2001). Genetic correlates of musical pitchrecognition in humans. Science 291, 1969-1972. doi:10.1126/science.291.5510.1969.
Freeman, W. (2000). "A neurobiological role of music insocial bonding," in The Origins ofMusic, eds. N.L. Wallin, B. Merker & S. Brown. (Cambridge, MA: MIT Press),411-424.
Hagen, E.H., and Bryant, G.A. (2003). Music and dance as acoalition signaling system. Human Nature14, 21-51. doi:10.1007/s12110-003-1015-z.
Koelsch, S. (2014). Brain correlates of music-evokedemotions. Nat. Rev. Neurosci. 15, 170-180. doi: 10.1038/nrn3666.
Koelsch, S., Fritz, T., Von Cramon, D.Y., Müller, K., andFriederici, A.D. (2006). Investigating emotion with music: An fMRI study. Hum. Brain Mapp. 27, 239-250. doi: 10.1002/hbm.20180.
Li, W.J., Yu, H., Yang, J.M., Gao, J., Jiang, H., Feng, M.,Zhao, Y.X., and Chen, Z.Y. (2010). Anxiolytic effect of music exposure onBDNF(Met/Met) transgenic mice. Brain Res.1347, 71-79. doi:10.1016/j.brainres.2010.05.080.
Menon, V., and Levitin, D.J. (2005). The rewards of musiclistening: Response and physiological connectivity of the mesolimbic system. Neuroimage 28, 175-184. doi: 10.1016/j.neuroimage.2005.05.053.
Miller, G. (2000). "Evolution of human music through sexualselection," in The Origins of Music,eds. N.L. Wallin, B. Merker & S. Brown. (Cambridge, MA: MIT Press), 329-360.
Mithen, S.J. (2007). "Music and the origin of modernhumans," in Rethinking the HumanRevolution, eds. P. Mellars, K. Boyle, O. Bar-Yosef & C. Stringer. (Cambridge: McDonald Institute), 107-117.
Patel, A.D., Iversen, J.R., Bregman, M.R., and Schulz, I.(2009). Experimental evidence for synchronization to a musical beat in anonhuman animal. Curr. Biol. 19, 827-830. doi: 10.1016/j.cub.2009.03.038.
Peretz, I., Champod, A.S., and Hyde, K. (2003). Varieties ofmusical disorders. Ann. N. Y. Acad. Sci.999, 58-75.
Peretz, I., Cummings, S., and Dubé, M.-P. (2007). Thegenetics of congenital amusia (tone deafness): A family-aggregation study. Am J Hum Genet. 81, 582-588. doi: 10.1086/521337.
Phillips-Silver, J., and Trainor, L.J. (2005). Feeling thebeat: movement influences infant rhythm perception. Science 308, 1430. doi:10.1126/science.1110922.
Roederer, J.G. (1984). The search for a survival value ofmusic. Music Perception, 350-356.
Schachner, A., Brady, T.F., Pepperberg, I.M., and Hauser,M.D. (2009). Spontaneous motor entrainment to music in multiple vocal mimickingspecies. Curr. Biol. 19, 831-836. doi:10.1016/j.cub.2009.03.061.
Sievers, B., Polansky, L., Casey, M., and Wheatley, T.(2013). Music and movement share a dynamic structure that supports universalexpressions of emotion. Proc. Natl. Acad.Sci. U. S. A. 110, 70-75. doi:10.1073/pnas.1209023110.
Tan, Y.T., Mcpherson, G.E., Peretz, I., Berkovic, S.F., andWilson, S.J. (2014). The genetic basis of music ability. Front. Psychol. 5. doi: 10.3389/fpsyg.2014.00658.
Trehub, S.E. (2003). The developmental origins of musicality.Nat. Neurosci. 6, 669-673. doi: 10.1038/nn1084.
Uchiyama, M., Jin, X., Zhang, Q., Amano, A., Watanabe, T.,and Niimi, M. (2012). Music exposure induced prolongation of cardiac allograftsurvival and generated regulatory CD4(+) cells in mice. Transplant. Proc. 44,1076-1079. doi: 10.1016/j.transproceed.2012.02.008.
Wieczorkowska, A., Synak, P., and Raś, Z.W. (2006). "Multi-label classification of emotionsin music," in Advances in SoftComputing, eds. M.A. Klopotek, S.T. Wierzchon & K. Trojanowski. (Berlin: Springer-Verlag Berlin), 307-315.
https://m.sciencenet.cn/blog-2481600-881815.html
上一篇:音乐起源之谜(三) ——音乐才能的遗传基础
下一篇:音乐起源之谜(五)——音乐之始,大道至简