博文
音乐起源之谜(六)——音乐和舞蹈统一于节奏运动
 精选
精选
||||
——音乐和舞蹈统一于节奏运动
前言:上一节对音乐的简化得出,一条时间线上的“节奏性相对音高”是旋律的本质,是音乐的本质。本节将通过声音的多普勒效应把“节奏性相对音高”解析为抽象发声物体的节奏性运动,从而对音乐认知中的“旋律不变性”问题给出解释,而且将音乐和舞蹈统一于“节奏运动”。进而将对音乐起源的探究,转化为对人类欣赏和制造节奏运动的研究。
纷繁复杂的音乐现象已经从音乐感知和认知层面简化到“节奏性相对音高(rhythmic relative pitches,RRPs)”,接下来需要从RRPs着手继续探究音乐的起源。一如我们在享用美食时不会去想食物里糖、脂、蛋白质等成分,以及它们的分子结构和营养效果;我们在欣赏音乐时也不会去想RRPs相关的问题,甚至绝大多数人根本没听过RRPs这个奇怪的东西;在这两种情况下,我们只会本能地去享受美食的香甜可口和音乐的优美曼妙。但是要研究音乐的起源,我们必须回答这个问题:人类为什么会欣赏RRPs?如前所述,我们的“眼、耳、鼻、舌、身”能感知外界的“色、声、香、味、触”等信息,我们耳朵感知到的声音,归根结底是一种信息载体。既然从音乐认知的层面已经不能再对RRPs进行简化,那么有必要从信息传递的角度研究RRPs所承载的更深层次内容。
声音及其信息传递功能
宇宙飞船的太阳能帆板接收光波是为了获得能量,虽然声波也能传递能量,显然动物的耳朵不是用来获取能量,而是用来获取信息的。和眼睛对可见光的感知一样,耳朵对声音的感知可以帮助动物了解环境中正在发生的事情,以便决策下一步的行为,最终获得生存和繁衍的利益。
故事里小动物听到木瓜掉到湖里的声音就四散而逃,“咕咚”声被误以为是怪兽;电影《大话西游》里唐僧那句“大雷啦,下雨啦,收衣服啦”戏谑而经典,道出了亘古不变的真理,“轰隆”的雷鸣往往预示着即将到来的暴雨;成语里“隔墙有耳”一词,寓意作为语言的人声承载着更加复杂的信息,小到家长里短,大到国事军情。由此可见,无论是最简单的“咕咚”,还是最复杂的语言,都承载着大量的信息。
以上是声音作为一个整体在传递信息中的作用,如果把声音解剖成部分,其不同要素也可以传递不同的信息。声音本身含有三个基本要素:响度(loudness)、音色(timbre)和音高(pitch)(Hubbard, 2010)。音色由发声物体的材料和质地等因素决定,所以音色可以鉴定发声物体或个体的身份。比如,我们可以靠声音区分钢琴和小提琴,区分张三李四和王五。声音的响度由发声物体的振幅决定,物体振幅越大,声音响度越大;反之,物体振幅越小,声音响度越小。不同的响度可以度量一个发声事件的大小。音高由发声物体的振动频率决定,振动频率越大,声音音高越高;反之,振动频率越小,声音音高越低。不同音高的声音也可以传递不同的信息,动物在遇到危险的时候往往会发出高音信号警示或救助于同类,比如一些女士遇到老鼠或蟑螂时的惊声尖叫。
现在回到音乐的最简模式RRPs。RRPs非但不是一个完整的声音信号,甚至连声音的基本要素都不算,因为RRPs不是一段连续的绝对音高,而是一段连续的相对音高。由于某些动物的鸣叫和人类的语言也是一段连续的声音组合,于是有科学家提出音乐是语言进化的副产品。如果按这个说法,RRPs应该和语言有莫大联系。不幸得很,虽然世界上语言种类众多,但是调性语言甚少。作为调性语言的汉语,虽然使用者几乎占全球四分之一人口,但是汉语里“妈、麻、马、骂”的调性和音乐里的RRPs也完全不相干;因为我们理解每句话的含义,不是基于这句话里每个字的声调组合,更不是基于每个字声调之间的相对音高,而是基于每个字的意义组合。
RRPs到底是何方神圣?到底传递了何种信息?其传递的信息对动物和人类的生存繁衍有何意义?上面这些讨论算是排除干扰,下面直入要害。
多普勒效应及回声定位系统
1842年,奥地利物理学家及数学家Christian Doppler提出了“多普勒效应(Doppler effect)”。根据多普勒效应,光波、声波、水波等辐射源与观察者之间的相对运动,会影响这些辐射波的观测频率 (Rosen, 2009)。当辐射源与观察者之间距离缩小时,相关波的观测频率增加,称为“蓝移(blue shift)”;反之,当辐射源与观察者之间距离增加时,相关波的观测频率减小,称为“红移(red shift)”。光波的多普勒效应一般应用于天体学,水波的多普勒效应可以在行进着船只和游动着天鹅的湖面观察到,而声音的多普勒效应可以在汽车和飞机的行进中观察到(图1)。

图1. 多普勒效应。A. 天体的蓝移和红移(示意图)与光波的多普勒效应相关(图片来源:网络);B. 水波的多普勒效应(图片来源:维基百科Doppler effect词条);C. 声波的多普勒效应(图片来源:网络)。
相较光波和水波的多普勒效应,声音的多普勒效应更贴近自然和生活。某些动物,比如一些蝙蝠、鲸鱼和海豚(图2,A)具有回声定位系统 (Au and Simmons, 2007; Parker et al., 2013)。这些动物通过发出声音脉冲信号、接收和分析回声信号,以探测周遭事物及其运动状态 (Nelson and MacIver, 2006)。蝙蝠在黑夜中不但能探测到周围的障碍,从而有效避闪;而且能探测到猎物的运动方向及速度,从而成功捕获猎物 (Ulanovsky and Moss, 2008; Corcoran and Conner, 2014)。基于动物回声定位系统的原理,人类发展出了声呐系统,声呐系统被装备于船只和潜艇,用于鱼群探测、海底测绘、敌情侦察等(图2,A)。
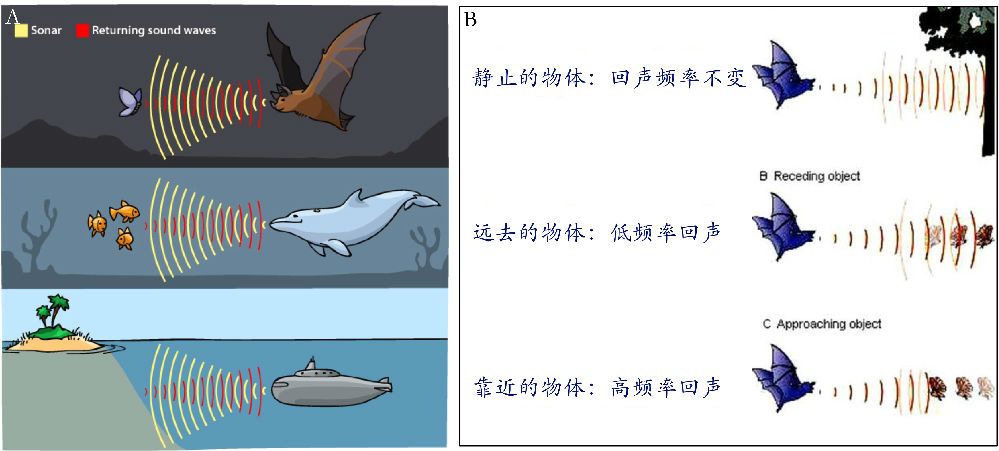
图2. 回声定位系统和多普勒效应。某些蝙蝠和鲸鱼利用回声定位系统探测和跟踪猎物,潜艇等航海设备所装备的声呐系统依此设计(A);回声定位系统和声呐系统对猎物和外界环境的探测,涉及多普勒效应的应用(B)。图片来源:网络。
不管是动物天生的回声定位系统,还是人造的声呐系统,都会涉及到对声波多普勒效应的分析(表1)。以蝙蝠为例(图2,B;图3,A),对猎物运动状态的明察秋毫是成功捕猎的前提条件之一,不过猎物的运动状态对蝙蝠来说是一个未知数,蝙蝠需要用一整套的感觉运动器官和神经系统来求解之。首先,蝙蝠已知自己发出的超声波频率,以及自身的运动状态(包括运动的方向和速度);然后,当超声波遇到环境中静止或运动的物体后,反射回来被蝙蝠接收,这样又多了一个已知条件;最后,蝙蝠的大脑整合上述已知条件,计算出障碍物的形状和距离,以及猎物的运动方向和速度。
由此可见,某些动物利用多普勒效应将声音频率的变化解析为猎物的运动状态。巧的是,RRPs也是一系列变化的声音频率,那么RRPs和多普勒效应是否有联系呢?
表1. 多普勒效应中观察频率f与发射频率f0之间的关系 (Wang, 2015)

大脑将“节奏性相对音高”解析为“节奏运动”
虽然人类不具备蝙蝠和鲸鱼那样高超的回声定位系统,不过回声定位功能在人类并不罕见。Stroffregen等人早在20年前就综合前人研究,提出回声定位可能是人类最基本的感知能力之一 (Stroffregenand Pittenger, 1995)。近年的研究表明,无论是盲人还是正常视力的人群,都具有或多或少的回声定位能力 (Schenkman and Nilsson, 2010),人们通过舌头或指关节制造声响,分析这些声响遇到障碍后发出的回声,以此对外部环境达到一定的认知 (Buckingham et al., 2014; Fiehler et al., 2015)。
即便人类的回声定位能力远逊某些动物,我仍然认为人类保有发达机制以解析声音的多普勒效应,其进化意义在于认知环境中声音事件的运动状态。据此我提出假说:人类在欣赏音乐时,将音乐中的“节奏性相对音高(RRPs)”转化成了一个虚拟发声物体的“节奏运动”(图3)。人们对音乐旋律的欣赏,就是对节奏运动的欣赏 (Wang, 2015)。
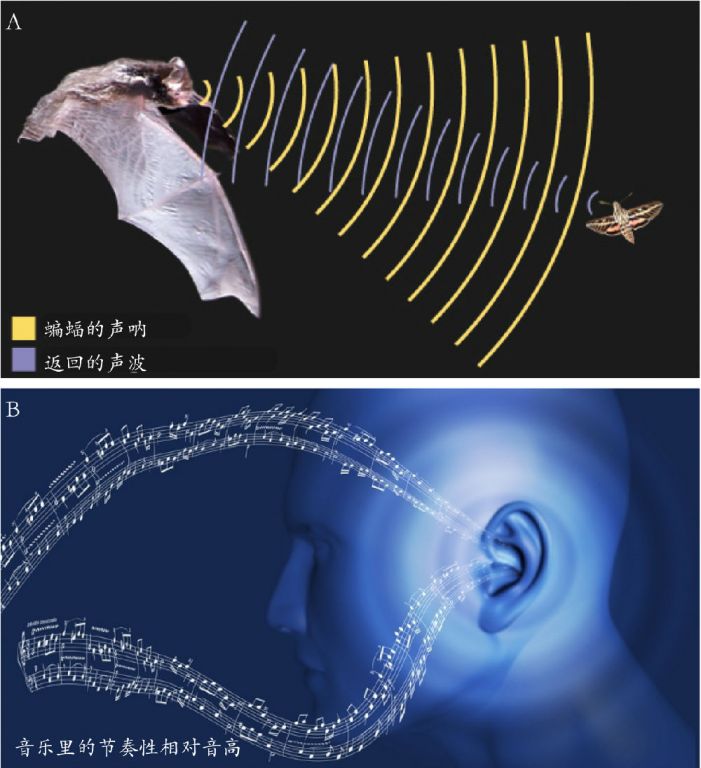
图3. 人脑将“节奏性相对音高”解析为“节奏运动”。即便人类的回声定位能力远逊某些动物,我仍然认为人类保有发达机制解析声音的多普勒效应。并据此提出假说认为:人类在听音乐时,将音乐中的“节奏性相对音高”转化成了一个虚拟发声物体的“节奏运动” (Wang,2015)。图片来源:网络。
这个提法虽然新颖而大胆,不过同时也面临一些问题,主要可能有三方面,我逐一阐述如下。
第一个问题:人脑将音乐旋律转化为节奏运动是个重大的事件,为什么我们不知道?人们对自己的认知可以分为四种:我知道我知道,我知道我不知道,我不知道我知道,我不知道我不知道。我想,我们对人脑将音乐旋律转化为节奏运动的认识应该属于“我不知道我知道”。大脑一直在将音乐旋律转化为节奏运动,这是“我知道”;但是我们在日常生活中根本没有意识到这个事件,所以属于“我不知道‘我知道’”。换句话说,我不知道我的大脑知道音乐里蕴含的节奏运动。然而事实上,对于大脑将音乐解析为运动这件事情,我们并不是完全不知道。在听音乐时,我们总感觉有个声音在跳动,常常会联想起一些运动的事件,比如轻柔的音乐旋律可能让我们想到微风拂柳,而激烈的旋律可能会让我们想到战争和体育运动。所以,对于人脑将音乐旋律转化为节奏运动这件事情,我们只是未能意识到,或者略微意识到。
第二个问题:现实中发声物体的节奏运动是否可以通过多普勒效应变成音乐?这个问题曾经一度困扰我,甚至想要放弃这个假说。图4是一个虚拟的系统:假设在一条足够长的笔直铁轨上,有一列足够长的火车,火车上有一辆可以在火车顶前后运动的汽车,汽车上载有一个可以持续发声的小号。理论上,当小号持续发出一个音高不变的声音(这里姑且称之“初音”),随着汽车的前后运动,站在离铁轨足够远处A点的人,可以因为多普勒效应听到类似音乐的声音。

图4. 以节奏运动“演奏”音乐的虚拟系统。假设在一条足够长的笔直铁轨上,有一列足够长的火车,火车上有一辆可以在火车顶前后运动的汽车,汽车上载有一个可以持续发声的小号。理论上,当小号持续发出一个音高不变的声音,随着汽车的前后运动,站在离铁轨足够远处A点的人,可以因为多普勒效应听到类似音乐的声音。图片来源:网络,经过PS。
这个虚拟系统貌似可以在现实中用发声物体的节奏运动“演奏”音乐,事实上有两个缺陷。其一,这个系统只能“演奏”比“初音”高一个八度和低两个八度之间的音,因为汽车的速度一旦突破音速340m/s (表2),这个系统就失效了。比如《沧海一声笑》(图5),小号的初始音可以是中音1、2、3、5、6中的任意一个,即便以中音6开始,汽车只需要以170m/s的速度背向听者行驶,就能“演奏”低音6这个音,再大一点速度可以“演奏”低音5,所以理论上来讲,通过控制汽车的行驶状态可以完整地“演奏”这首曲子;但是初始音不能是低音5或6,如果以低音5或6开始,小汽车只能以接近音速面向A点行驶,A点的听者才能听到接近中音5或6的两个音,小汽车根本不能完整地“演奏”这首曲子。其二,在这个系统中发声物体的节奏和听者听到的节奏不同步,因为小车偏离原点以后,小号与听者之间的距离会发生变化,而声音的传播会因为距离的变化而提前或延后到达听者。如果汽车的运动保持严格的节奏,那么A点听到的“音乐”节奏会丧失严格性;反之,如果想要保证“音乐”具有严格的节奏,那么汽车的运动节奏会丧失严格性。
表2. 音乐中固定的相对音高可以通过多普勒效应解释为不变的运动模式 (Wang, 2015)
图5. 《沧海一声笑》的简谱。图片来源:网络。
上述两个缺陷,使得在现实生活中无法重现节奏运动对音乐的“演奏”,似乎对“音乐旋律体现节奏运动”这一假说构成巨大挑战。然而现实生活中不存在或不能重建的事物,并不见得在认知水平也不存在。人类大脑通过多普勒效应对音乐节奏的解析机制,或许并不必须受声音在空气中传播速度的限制。我认为,发自乐器或播放器的定点RRPs,恰恰突破了上述两个限制,让人类可以欣赏到更加广阔和纯粹的节奏运动。
第三个问题:人类大脑是否存在将音乐旋律解析为节奏运动的神经和基因基础?这个问题最关键!如果说人类大脑具有将音乐旋律解析为节奏运动的机制,那么应该有相应的神经环路和基因基础为这一机制负责。遗憾的是目前还没有相关研究,所以这个问题悬而未决,对于这个问题的研究需要认知、神经、生物等学科的交叉。我先把问题摆在这里,希望能抛砖引玉,期待以后的研究能证明或证伪之。
由此可见,前两个问题不必太过担忧,第三个问题有待继续研究。虽然目前在生物学水平上没有直接的证据,不过该假说对“旋律不变性”的解释、对音乐和舞蹈的统一,可以作为间接证据;当然,这些间接证据的证明力度是有限的。
“旋律不变性”体现“运动模式不变性”
上一节《音乐之始,大道至简》里,我有提到关于音乐认知的“旋律不变性”问题。所谓的“旋律不变性”,是指一个既定的旋律,不管它以哪个绝对音高开始演奏,只要其随后一系列音之间的相对音高保持不变,我们就认为它是同一个旋律 (McDermott and Oxenham, 2008; Cariani and Micheyl, 2012; Trainor and Unrau, 2012)。言下之意,旋律中的“节奏性相对音高(RRPs)”才是音乐的万变不离之“宗”。
本节引入声音的多普勒效应,认为人类大脑将RRPs解析为一个虚拟发声物体的节奏运动。回到图4那个虚拟系统,以汽车的节奏运动“演奏”《沧海一声笑》,如果小号以中音1、2、3、5、6中的任何一个为初音,汽车的运动模式,包括速度、方向和路径都必须保持一致;当小号以固定的初音,比如中音6“演奏”,在汽车运动模式不变的情况下,通过调整火车的运动模式就可能“演奏”出比原来低八度、高八度、甚至变调的《沧海一声笑》。我们可以发现,不管哪种情况,汽车的运动模式都是相对恒定的。
可见,音乐中的任何既定旋律,不管以低八度音、高八度音,或者变调演奏,在大脑里都有且只有一个既定的运动模式与之对应。所以,该假说可以解释音乐认知领域的“旋律不变性”问题。
音乐和舞蹈统一于节奏运动
音乐和舞蹈像一对双胞胎,常常如影随形。即便只是单纯的聆听音乐,人类 (Repp, 2005)以及某些动物 (Patel et al.,2009; Schachner et al., 2009; Cook et al.,2013)也会随着音乐节奏点头、晃腿、叩动手指等。此外,音乐认知也和身体运动紧密联系 (Phillips-Silver and Trainor, 2005;Zatorre et al., 2007; Trainor et al., 2009; Maes et al., 2014)。
旋律和舞蹈分别是排列在一条时间线上的音高和动作。既然大脑将旋律解析为节奏运动,那么音乐和舞蹈可以统一成一个共同模式:节奏运动(图6)。不过舞蹈的节奏运动是具体的,而音乐的节奏运动是抽象的,这两类节奏运动具有不同的感知途径,却在更高的认知层面达成统一,可谓殊途同归。据此,音乐和舞蹈在行为和认识水平的紧密联系可以得到解释。
音乐和舞蹈的节奏运动被人类感知(perception)、制造(production)和同步化(synchronization),在我们大脑中诱导出快感和其它正面或负面情绪。那么接下来的问题是,为什么节奏运动能够诱导这些快感和情绪?
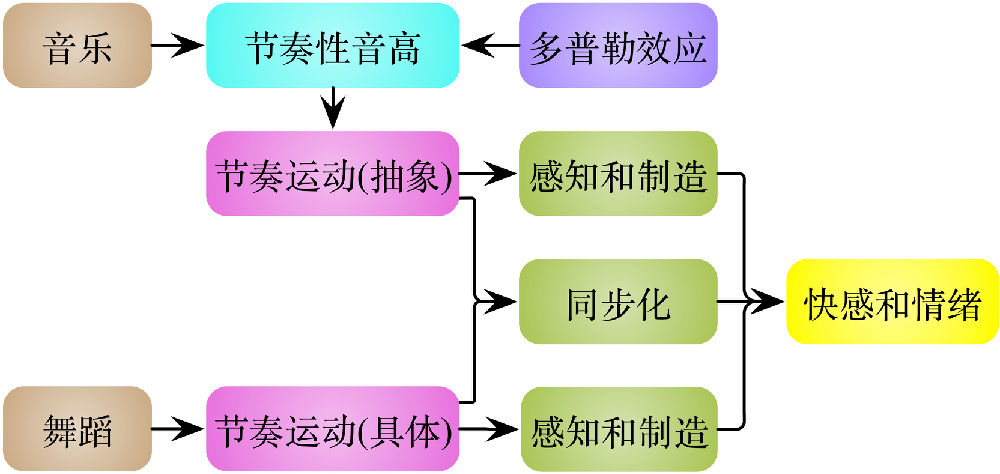
图6.音乐和舞蹈统一于“节奏运动” (Wang, 2015)。旋律和舞蹈分别是排列在一条时间线上的音高和动作,通过多普勒效应,音乐和舞蹈可以得到统一。对这些节奏运动的感知、制造和同步化,在大脑中诱导出快感和其它正面或负面情绪。
注:1.该博文中凡有确切来源图片均已注明出处,其它图片收集自网络,原作者可以联系我撤销图片或添加备注。
2.成文仓促,难免失误,请大家多多指正;另,有交流或合作意向的老师和同学,欢迎随时联系。
王天燕(tianyanwang@hotmail.com)
2015年05月08日
相关链接:
A hypothesis on the biological origins and social evolution of music and dance (Wang, Front. Neurosci. 2015)
音乐起源之谜(一)——音乐、舞蹈和语言起源的“节奏适应假说” (王天燕,03/19/2015)
音乐起源之谜(二)——音乐的“文化起源” VS“生物起源” (王天燕,03/26/2015)
音乐起源之谜(三)——音乐才能的遗传基础 (王天燕,04/02/2015)
音乐起源之谜(四)——音乐起源的性选择假说及其不足(王天燕,04/12/2015)
音乐起源之谜(五)——音乐之始,大道至简 (王天燕,04/16/2015)
音乐起源之谜(六)——音乐和舞蹈统一于节奏运动 (王天燕,05/08/2015)
音乐起源之谜(七)——节奏适应是食物和性的前提 (王天燕,05/15/2015)
音乐起源之谜(八)——音乐在动物界的起源和进化 (王天燕,05/22/2015)
音乐起源之谜(九)——人类音乐的起源和进化 (王天燕,06/08/2015)
音乐起源之谜(十/完)——节奏适应假说的机遇和挑战 (王天燕,06/18/2015)
参考文献:
Au, W.W.L., and Simmons,J.A. (2007). Echolocation in dolphins and bats. Physics Today 60, 40-45.doi: 10.1063/1.2784683.
Buckingham, G., Milne, J.L., Byrne, C.M., and Goodale, M.A. (2014). TheSize-Weight Illusion Induced Through Human Echolocation. Psychological science,0956797614561267.
Cariani, P., and Micheyl, C. (2012). "Toward a theory of informationprocessing in auditory cortex," in TheHuman Auditory Cortex, eds. D. Poeppel, T. Overath, A.N. Popper & R.R.Fay. (New York, NY: Springer), 351-390.
Cook, P., Rouse, A., Wilson, M., and Reichmuth, C. (2013). A Californiasea lion (Zalophus californianus) can keep the beat: motor entrainment torhythmic auditory stimuli in a non vocal mimic. Journal of Comparative Psychology 127, 412-427. doi: 10.1037/a0032345.
Corcoran, A.J., and Conner, W.E. (2014). Bats jamming bats: foodcompetition through sonar interference. Science346, 745-747. doi:10.1126/science.1259512.
Fiehler, K., Schütz, I., Meller, T., and Thaler, L. (2015). NeuralCorrelates of Human Echolocation of Path Direction During Walking. Multisensory Research 28, 195-226.
Hubbard, T.L. (2010). Auditory imagery: empirical findings. Psychological bulletin 136, 302.
Maes, P.-J., Leman, M., Palmer, C., and Wanderley, M.M. (2014).Action-based effects on music perception. Frontiersin Psychology 4. doi: 10.3389/fpsyg.2013.01008.
Mcdermott, J.H., and Oxenham, A.J. (2008). Music perception, pitch, andthe auditory system. Current Opinion inNeurobiology 18, 452-463. doi:10.1016/j.conb.2008.09.005.
Nelson, M.E., and Maciver, M.A. (2006). Sensory acquisition in activesensing systems. Journal of ComparativePhysiology. A, Sensory, Neural, and Behavioral Physiology 192, 573-586. doi:10.1007/s00359-006-0099-4.
Parker, J., Tsagkogeorga, G., Cotton, J.A., Liu, Y., Provero, P., Stupka,E., et al. (2013). Genome-wide signatures of convergent evolution inecholocating mammals. Nature 502, 228-231. doi: 10.1038/nature12511.
Patel, A.D., Iversen, J.R., Bregman, M.R., and Schulz, I. (2009).Experimental evidence for synchronization to a musical beat in a nonhumananimal. Current Biology 19, 827-830. doi:10.1016/j.cub.2009.03.038.
Phillips-Silver, J., and Trainor, L.J. (2005). Feeling the beat: movementinfluences infant rhythm perception. Science308, 1430. doi:10.1126/science.1110922.
Repp, B.H. (2005). Sensorimotor synchronization: A review of the tappingliterature. Psychonomic Bulletin &Review 12, 969-992. doi:10.3758/bf03206433.
Rosen, J.G., Lisa Quinn (2009). Encyclopediaof Physical Science. New York, NY: Facts on File.
Schachner, A., Brady, T.F., Pepperberg, I.M., and Hauser, M.D. (2009).Spontaneous motor entrainment to music in multiple vocal mimicking species. Current Biology 19, 831-836. doi: 10.1016/j.cub.2009.03.061.
Schenkman, B.N., and Nilsson, M.E. (2010). Human echolocation: blind andsighted persons’ ability to detect sounds recorded in the presence of areflecting object. Perception 39, 483.
Stroffregen, T.A., and Pittenger, J.B. (1995). Human echolocation as a basicform of perception and action. Ecologicalpsychology 7, 181-216.
Trainor, L.J., Gao, X., Lei, J.-J., Lehtovaara, K., and Harris, L.R.(2009). The primal role of the vestibular system in determining musical rhythm.Cortex 45, 35-43. doi: 10.1016/j.cortex.2007.10.014.
Trainor, L.J., and Unrau, A. (2012). "Development of pitch and musicperception," in Human AuditoryDevelopment, eds. L. Werner, R.R. Fay & A.N. Popper. (New York, NY: Springer), 223-254.
Ulanovsky, N., and Moss, C.F. (2008). What the bat's voice tells the bat'sbrain. Proceedings of the NationalAcademy of Sciences of the United States of America 105, 8491-8498. doi: 10.1073/pnas.0703550105.
Wang, T. (2015). A hypothesis on the biological origins and socialevolution of music and dance. Front.Neurosci. 9. doi: 10.3389/fnins.2015.00030.
Zatorre, R.J., Chen, J.L., and Penhune, V.B. (2007). When the brain playsmusic: auditory–motor interactions in music perception and production. Nature Reviews Neuroscience 8, 547-558. doi: 10.1038/nrn2152.
https://m.sciencenet.cn/blog-2481600-888460.html
上一篇:音乐起源之谜(五)——音乐之始,大道至简
下一篇:音乐起源之谜(七)——节奏适应是食物和性的前提