博文
[转载]来自徐凌燕科学网博客:代谢学人--代谢典藏 |Nature综述:脂肪代谢全攻略 (下篇)
||
代谢学人--代谢典藏 |Nature综述:脂肪代谢全攻略 (下篇)
||
Nature Reviews Endocrinology 综述
代谢典藏 |Nature综述:脂肪代谢全攻略 (下篇)
撰文 | 许赛男 陈俊桐 王赛男 胡天惠 邱瑾
编辑 | 孟美瑶

经过上期的讲解
大家是不是对白色脂肪有了一定的了解呢?
错过精彩的朋友看这里↓↓↓↓↓↓

接下来!
挺直腰板!
继续学习白色脂肪的代谢过程吧!
能量消耗
哺乳动物细胞有多种机制消耗能量,其中线粒体氧化呼吸是产热的主要方式。底物循环,有时被不恰当地称为无用循环,也有助于能量消耗,并作为调控新陈代谢的关键步骤被其他综述总结。棕色和米色脂肪细胞有着非常高的脂肪酸氧化速率。与白色脂肪细胞相比,产热脂肪细胞具有独特的功能,可以将能量以热的形式散失而非以化学形式储存。有证据表明,产热脂肪细胞是从祖细胞从头分化而来。早期研究表明,在冷刺激或β3-肾上腺素能和PPAR激动剂处理后,白色脂肪细胞可以直接转化为米色脂肪细胞。在本篇综述中,我们着重关注单房白色脂肪细胞与米色脂肪细胞的相互转化。
米色脂肪细胞典型的适应性产热途径涉及到解偶联蛋白1(UCP1)。UCP1位于线粒体内膜,它可以消散线粒体内外膜间的质子电化学梯度,是这些质子势能不能被用于生成ATP(图5a)。UCP1隶属于组分众多的、能够以热量形式散失能量的细胞机器的一员。在人类脂肪细胞由白色转变为米色的过程中,线粒体持续分裂导致线粒体碎片化,从而使得UCP1依赖的呼吸解偶联增强。同时,该转变过程引起显著的代谢重编程,在细胞质和线粒体中促进脂肪酸合成和分解代谢。丙酮酸脱氢酶复合体被丙酮酸脱氢酶激酶4磷酸化,从而抑制其活性,将葡萄糖利用从氧化途径重定向到TAG合成,从而促使解偶联状态下的线粒体转而利用脂肪酸作为主要能量来源。除了通过肾上腺素能信号通路调控UCP1的表达和活性外,琥珀酸作为TCA循环的中间产物,也可以通过刺激棕色和米色脂肪中活性氧的生成来激活UCP1介导的产热过程。此外,胞外的琥珀酸很容易被棕色脂肪细胞吸收并氧化,但人类米色脂肪细胞如何响应胞外琥珀酸,以及在体育锻炼和缺血等促琥珀酸释放的条件下,琥珀酸是否能激活产热,仍不清楚。

图5:脂肪细胞中的能量消散。
a.依赖于UCP1的生热作用涉及脂肪酸β-氧化和琥珀酸氧化诱导的ROS(活性氧)生成。响应产热刺激,脂肪酸、葡萄糖和琥珀酸从循环中被摄取。脂肪酸也可以从细胞内储存的TAG中被动员。脂肪酸相比于葡萄糖被优先氧化利用,这是由于PDH(丙酮酸脱氢酶)复合体组分被PDK4(丙酮酸脱氢酶激酶4)磷酸化而受到抑制。
b.不依赖于UCP1的多种能量消散途径。TAG合成和水解途径中的底物循环,包括脂解和甘油被磷酸化形成成甘油-3P(甘油-3-磷酸),从而使脂肪酸再酯化。为这个循环提供能量的另一个分支是从葡萄糖合成脂肪酸的途径(脂质从头合成)。TAG-脂肪酸底物循环对产热的贡献是有争议的。内质网中的SERCA2-兰尼定受体(RyR)通路诱导Ca2+循环。在线粒体中,肌酸底物循环也能够产生能量消散。这后两种途径同时伴随着ATP合成、消耗和热量产生。ETC,电子传递链;IMS,线粒体膜间隙;RCC,呼吸链复合物;S, F1/F0,ATP合成酶;TCA,三羧酸循环。
相反的,在热中性环境下的小鼠,以及小鼠和人类的衰老过程中,都观察到脂肪细胞由棕色向白色转化。向白色转化的棕色脂肪细胞具有单房脂滴这一经典的白色脂肪细胞特征,但其线粒体仍含有较低含量的UCP1,并保留了棕色脂肪细胞样特征。与肥大和功能障碍的白色脂肪细胞相同,变白的棕色脂肪细胞中炎性小体被激活,并能够促使细胞焦亡。在温暖情况下,米色脂肪细胞经历染色质重塑向白色状态转化,但在棕色脂肪细胞中未观察到这一现象。然而,米色脂肪细胞保留了表观遗传记忆,当其再次暴露在寒冷中时,会重新激活产热程序。这些数据表明白色和米色脂肪细胞可以双向转化,棕色脂肪组织中的单房白色样脂肪细胞可能是潜在的棕色脂肪细胞。特定的染色质重塑酶介导了这种转化。组蛋白甲基化在化学上非常稳定的,因此可以作为一种长期的细胞记忆机制。有多种调节组蛋白赖氨酸残基甲基化的酶,特别是赖氨酸特异性脱甲基酶(KDM)家族成员,参与米色脂肪细胞的代谢调控。其中,研究最多的家族成员是KDM1A,它通过与产热转录因子ZFP516和PRDM16相互作用,激活米色脂肪基因程序而抑制白色脂肪特异基因。总之,对于多种酶的研究表明,组蛋白H3赖氨酸残基的去甲基化对于维持米色脂肪细胞表型至关重要。
近年来,在米色和白色脂肪细胞代谢中,发现了多个不依赖于UCP1的产热过程(图5b)。TAG-脂肪酸循环是脂肪细胞中公认的底物循环。在脂解过程中,脂肪酶分解TAG产生脂肪酸和甘油,这些产物随后在脂肪细胞中被输出或被氧化。甘油在甘油激酶的作用下磷酸化以及脂肪酸活化形成脂酰辅酶A,使得TAG能够被再酯化。甘油激酶在人类白色脂肪细胞中的表达和活性远低于棕色和米色脂肪细胞。但是肾上腺素能激活和PPAR激动剂处理可以诱导白色脂肪细胞中甘油激酶的活性,从而精密调控脂肪酸的去向(释放、氧化和酯化)。由此可见,TAG-脂肪酸循环耗能很低。然而,当底物循环与其他途径如脂质从头合成和脂肪酸氧化相结合,典型的白色脂肪可能对能量消耗产生很大贡献,并促进小鼠的瘦表型。在恶性高热中,骨骼肌细胞肌浆网内Ca2+不受控制地释放,导致代谢亢进和热量生成。与此类似,当肌浆网/内质网Ca2+-ATP酶和兰尼定(Ryanodine)受体调控的ATP依赖性Ca2+循环增加时,米色脂肪可能发生冷诱导的产热现象。ATP依赖性Ca2+循环的重要功能在UCP1敲除的家猪米色脂肪细胞中被证实。此外,有证据表明,当米色脂肪中ADP受限时,线粒体底物循环受到肌酸的调节以驱动产热呼吸。无论UCP1依赖的质子泄漏是否发生,米色脂肪细胞中都发生这一无用肌酸循环,这种循环对细胞基础代谢率有显著贡献,而不是仅在受到急性肾上腺素能刺激时发挥作用。脂肪细胞特异性的肌酸转运减少会导致肾上腺素能诱导的产热功能受损。在人皮下脂肪细胞中,肌酸转运蛋白的表达与肥胖和胰岛素抵抗呈负相关。因此,这些数据表明细胞外肌酸可以调控米色脂肪介导的能量消耗。另一个皮下WAT中UCP1非依赖性生热的例子是AMP激酶基因激活小鼠中观察到的能量消耗增加,但其涉及的分子途径尚未确定。
因此,从新陈代谢的角度来看,脂肪细胞的类别为其中使用不同底物的代谢机器所决定。
代谢通路的相互交流
在白色脂肪组织中,脂质分解和生成通常被视为不同结局的两条独立途径。然而,小鼠在接受慢性肾上腺素能刺激后,不但促进了WAT中TAG的水解,也同时促进了DNL和脂质周转。有趣的是,Pnpla2敲除不仅如预期般减少了脂解,由于DNL酶的下调,也同时导致了脂质周转的减少(图6)。肾上腺素能刺激后,AGTL介导的和/或脂解介导的DNL激活的确切分子机制及其在人类WAT中的相关性尚不清楚。代谢通路相互交流的另一个例子是在脂肪细胞中高度表达的中性脂肪酶HSL。HSL部分敲除可改善肥胖小鼠的全身胰岛素敏感性,但血浆中游离脂肪酸水平未发生变化,表明其机制可能与脂解作用无关。在人源脂肪细胞和小鼠WAT中抑制HSL的表达也会导致胰岛素敏感性和DNL的增强,其中DNL酶极长链脂肪酸延长酶6(ELOVL6)表达上升最为显著。ELOVL6与硬脂酰去饱和酶一起促进磷脂油酸生成,从而增加质膜的流动性并增强胰岛素信号。在脂肪细胞中,ELOVL6是葡萄糖应答转录因子ChREBP(由MLXIPL编码)的主要转录靶点。从机制上讲,HSL与ChREBPα发生相互作用,削弱了ChREBPα转位至细胞核的能力,从而阻断了ChREBPα介导的ChREBPβ(高转录活性异构体,由MLXIPL的可变转录起始点编码产生)的转录,而WAT中ChREBPβ的表达与全身胰岛素敏感性密切相关。葡萄糖代谢也与支链氨基酸(BCAAs,如亮氨酸、异亮氨酸和缬氨酸)的代谢有关。与BCAAs在衰老或恶病质等条件下促进蛋白质合成的有益作用相反,血液中BCAAs水平的升高与人类肥胖、胰岛素抵抗、T2DM和心血管疾病相关。孟德尔随机化分析显示BCAA代谢与T2DM疾病发生间存在因果关系。在一定程度上,循环BCAA水平的升高与WAT中分解酶的表达下降和由此导致的脂肪酸氧化降低有关。蛋白质分解代谢可能提供支持线粒体代谢和DNL的BCAAs。在体外,产生自BCAAs分解的乙酰-CoA可占小鼠和人类白色脂肪细胞中脂质生成乙酰-CoA池的三分之一。
此外,线粒体BCAA分解代谢与DNL之间也存在着一定的联系。在脂肪组织中,脂肪酸合成酶和肉碱乙酰转移酶的酶催化多功能性促使BCAAs合成单甲基支链脂肪酸,并能够被掺入TAG。这一途径在何种生理条件下发挥重要作用尚未确定。在禁食期间支链脂肪酸被动员,而高脂饮食减少了这种动员现象,尽管这一现象的作用和重要性尚不清楚。在另一方面,当小鼠脂肪细胞中葡萄糖转运增强时,其WAT中BCAAs相关代谢酶的表达降低,但葡萄糖代谢如何调节BCAA酶的表达仍不清楚。这些研究描述了脂肪细胞中葡萄糖、脂肪酸和氨基酸代谢相互作用的一些例子。利用高级系统生物学方法建立脂肪细胞的全基因组代谢模型,或许将有助于我们更好地理解代谢通路之间的相互作用。
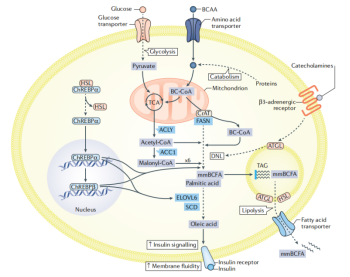
图6:白色脂肪组织中代谢通路的相互交流。HSL(激素敏感脂肪酶)和ChREBPα (碳水化合物反应元件结合蛋白α)之间的相互作用抑制ChREBPα进入细胞核。而抑制HSL-ChREBPα的相互作用能够促进ChREBPα的转录活性,进而诱导ChREBPβ(具有高转录活性的异构体)和脂质生成相关酶的表达,如ACLY(ATP柠檬酸裂解酶)、ACC1(乙酰辅酶a羧化酶)、FASN(脂肪酸合成酶)、ELOVL6(极长链脂肪酸延长酶6)和SCD (硬脂酰CoA去饱和酶)。ChREBP对于ELOVL6具有转录偏好性,ELOVL6能够促进油酸的合成并使其掺入质膜磷脂,从而增加质膜流动性,促进胰岛素的信号转导。与之相反,β3-肾上腺素能受体信号促进ATGL(脂肪甘油三酯脂肪酶)依赖的脂质生成酶表达和脂质从头生成(DNL)。支链氨基酸(BCAAs)是DNL另一重要底物来源。BCAAs可在TCA(三羧酸循环)中被氧化,为扩充脂质生成相关的乙酰-CoA库做出贡献。BCAA分解代谢的支链辅酶A中间体(BC-CoAs)在线粒体中产生,但可以通过CrAT(肉碱酰基转移酶)转运到胞浆。CrAT和FASN的酶催化多功能性有利于BC-CoA的延伸和mmBCFA(单甲基支链脂肪酸)的产生。mmBCFA可通过掺入三酰基甘油(TAG)储存并在禁食期间动员。葡萄糖代谢通过未知机制抑制BCAA代谢。
脂肪细胞代谢失衡
脂肪细胞脂滴紊乱。WAT过量或缺乏可能导致相似的病理表型。在这两种情况下,皮下WAT储存障碍都会导致脂质流向其他脂肪库和器官,如内脏脂肪、肝脏、骨骼肌和胰腺β细胞等。由此造成的脂质毒性导致这些器官的代谢功能改变,进而导致心血管代谢紊乱。其中,遗传性脂肪营养不良是具有临床意义的脂肪细胞代谢受损模型。疾病相关的AGPAT2和CAVIN1突变引起先天性全身性脂肪营养不良,这是一种罕见的常染色体隐性遗传病,其特征是脂肪组织几近完全缺乏(图3-4)。编码三种脂滴相关蛋白PLIN1、HSL和CIDEC的突变则引起家族性部分脂肪营养不良,这是一种常染色体隐性或常染色体显性疾病,以不同脂肪库中不同程度的体脂减少为特征。一般来说,脂肪减少的程度与其并发症的严重程度相关,如胰岛素抵抗、血脂异常、肝脏脂肪变性和多囊卵巢综合征。遗传学证据表明,人群中外周WAT储存多余脂质的能力受限与人类胰岛素抵抗(BOX2)有关,表明常见的遗传变异通过类似脂肪营养不良的机制影响心血管代谢疾病患病风险。值得注意的是,即使脂肪细胞大小增加,也可能无法形成或扩大脂肪库。总的来说,对人类和转基因小鼠模型的研究表明,脂肪营养不良和肥胖的代谢功能障碍是相似的。
肝脏葡萄糖生成。有力的证据表明,脂肪细胞中脂肪酸代谢紊乱能够影响整个机体。基础脂解率与胰岛素抵抗正相关,且独立于体重指数和年龄因素。在前瞻性队列中,基线时的高基础脂解率和低应激脂解率能够预测后期胰岛素抵抗的发生。来自脂肪酶缺陷小鼠模型和PLIN1缺陷患者的研究也表明脂肪细胞脂解作用对胰岛素抵抗的贡献。胰岛素介导的肝脏葡萄糖生成抑制受损是胰岛素抵抗的一大显著特征(图7)。胰岛素对肝脏葡萄糖生成的急性抑制与胰岛素诱导的WAT脂解抑制有关。由于向肝脏输送的脂肪酸减少,肝脏中丙酮酸羧化酶活性降低,因此降低了肝脏的乙酰辅酶A浓度和葡萄糖的生成。在高脂饮食引起的胰岛素抵抗的啮齿动物中,胰岛素抗脂解作用受损,从而促进肝脏葡萄糖生成。然而,在啮齿动物和实验狗中的研究结果表明,相对于肝外组织(如脂肪组织),胰岛素对肝细胞葡萄糖生成的直接调控作用占主导地位。胰岛素的直接或间接作用(即抗脂解作用)在调节肝脏葡萄糖生成中哪个更重要似乎取决于实验背景。尽管已经开展了大量的研究,但仍有几个问题没有得到解答。脂解产生的脂肪酸进入肝细胞后面临多种命运。随着生理病理状态的改变,脂肪酸对于不同代谢途径,尤其是TAG合成和脂肪酸氧化的相对贡献相应改变,并因此调节胰岛素对肝脏葡萄糖代谢的影响。脂解产生的脂肪酸的长期输出对于肥胖相关胰岛素抵抗发生发展的重要性在人类中仍缺乏有力证据。一项系统的文献总结显示,禁食条件循环中的NEFAs(非酯化脂肪酸)水平与人类体脂和胰岛素敏感性相关性很低。人体中,肝脏和WAT发生胰岛素抵抗的时间先后关系也没有被很好的研究。在肥胖个体中,这两种组织的胰岛素抵抗的发病时间可能不同。
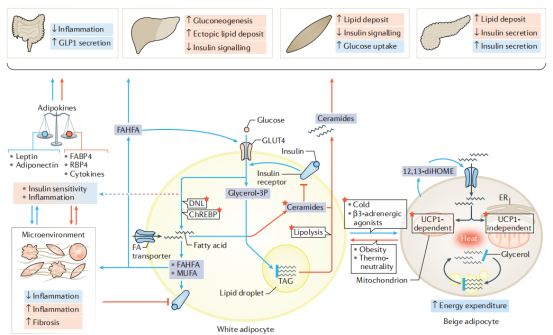
图7:脂肪细胞代谢的全身性影响和治疗前景。在非病理条件下,脂肪细胞的代谢是有益的,可以维持全身胰岛素敏感性(蓝色途径)。这种代谢表型促进脂滴以三酰甘油(TAG)形式健康的储存脂质,保持了较低的基础脂解作用、脂肪从头合成,并分泌有益的生物活性脂质(如羟基脂肪酸酯FAHFAs和12,13-diHOME)和脂肪因子(如瘦素和脂联素)的分泌。在肥胖或脂肪营养不良状态下,脂肪细胞表现出相反的特征,其分泌因子(如脂肪酸、神经酰胺、细胞因子和脂肪酸结合蛋白FABP4及视黄醇结合蛋白RBP4)促进慢性炎症和全身胰岛素抵抗(红色途径)。脂肪细胞代谢的多个方面都是药物开发的有价值的潜在治疗靶点(红色星号)。通过抑制HSL(激素敏感脂肪酶)与 ChREBP(碳水化合物反应元件结合蛋白)之间的相互作用,能够提高质膜磷脂中MUFAs(单不饱和脂肪酸)的含量,进而激活DNL和增强胰岛素信号。DNL的增强有助于有益脂质的合成,如 FAHFAs,从而提高全身胰岛素敏感性。抑制神经酰胺合成和/或激活神经酰胺降解对全身胰岛素抵抗有保护作用。Gαi-偶联GPCR激动剂能够抑制脂解,因此是另一种很有潜力的策略。PPARγ(过氧化物酶体增殖剂激活受体-γ)激动剂可以促进健康的脂质储存和DNL,减少炎症反应,诱导有益脂肪因子的分泌,改善全身胰岛素敏感性。最后,将白色脂肪细胞转化为米色脂肪细胞,并刺激米色脂肪细胞的能量耗散,是增加能量消耗的一种有吸引力的策略。ER:内质网;GLP1:高血糖素样肽1;glycerol-3P:甘油-3-磷酸;UCP1:解偶联蛋白1。
肿瘤侵袭性和恶病质。脂肪细胞的脂质分解和白色脂肪代谢除了影响糖尿病和心血管风险外,还影响某些癌症的发展和癌症相关恶病质。在乳腺癌中,肿瘤细胞的活跃分泌活性促使肿瘤周围脂肪细胞中脂质的消耗并大量释放脂肪酸。乳腺癌细胞对这些脂肪酸的摄取引起肿瘤细胞广泛的代谢重塑,从而增强肿瘤的侵袭性。癌症相关的恶病质会危及生命,在这一病理情况下,脂肪的减少可能先于瘦肉的减少。在一些癌症相关恶病质患者和动物模型中观察到应激脂解率升高,以及循环NEFA和甘油水平的增加。在荷瘤ATGL敲除小鼠模型中,观察到体重、WAT和骨骼肌质量流失的缓解现象。通过阻断CIDEA和AMP激酶(细胞能量稳态调节因子)之间的相互作用抑制白色脂肪细胞的脂解和DNL,能够部分抵抗脂肪流失,并延长荷瘤小鼠对恶病质的抵抗时长。有可能脂肪酸生成的增加促进了骨骼肌中脂类和脂毒性物质的积累,从而参与肌肉萎缩的发展。
脂肪酸对巨噬细胞的影响。白色脂肪源性脂肪酸已成为巨噬细胞代谢的重要调节剂。在脂解长期激活条件下,脂肪细胞释放的脂肪酸被巨噬细胞吸收并导致脂质累积。这种清除脂肪酸的作用与泡沫细胞在动脉粥样硬化斑块中积累胆固醇酯的情况类似。在小鼠模型中,肥胖与巨噬细胞中脂滴积累和溶酶体生物发生激活相关,从而导致TAG分解代谢。另外,如前所述,脂肪组织巨噬细胞中的脂质也可能来源于脂肪细胞释放的脂滴衍生的外泌体样囊泡。因此,WAT中脂肪酸的净释放收到脂肪细胞脂质动员和脂肪组织巨噬细胞溶酶体活性的共同调控。在人体中,脂肪细胞和巨噬细胞对于WAT脂肪酸释放的重要性目前尚不清楚。这些缓冲机制是否切实影响循环中的脂肪酸水平,从而影响这些脂肪酸在其他器官中以脂质形式异位累积,并减轻WAT脂解加强所带来的不利影响,仍有待研究。
早期研究表明,脂肪细胞释放的脂肪酸可能对脂肪组织中其他类型细胞产生影响,从而间接影响全身代谢。WAT中脂肪细胞与其他类型细胞之间的相互作用能够调节免疫应答和纤维化(BOX1,图7)。这种组织重塑改变了间质血管细胞的分泌表型,产生可能具有内分泌作用的分子。肥大的脂肪细胞在严重代谢应激下易发生焦亡,这是一种促炎性的程序性细胞死亡形式。肥胖WAT中死亡的脂肪细胞数量增加,从而招募巨噬细胞形成冠状结构。被招募来的巨噬细胞表现为促炎M1样表型,并产生一系列细胞因子和趋化因子。大量研究表明,脂肪组织中这一被称为低度炎症的表型对于促进肥胖和胰岛素抵抗发展具有显著作用。然而,在肥胖和禁食期间,人类和小鼠的WAT巨噬细胞的促炎M1样表型和这些细胞的脂质转运能力是否有重要作用尚不清楚,并且研究提示,可能随着脂肪库的解剖位置不同而有不同作用。尽管脂肪组织巨噬细胞具有以上这些特征,但本文总结的研究表明,脂肪细胞代谢可能仍是WAT免疫反应的主要驱动因素。例如,对具有不同程度肥胖和代谢障碍的女性的WAT转录组分析显示,皮下和内脏脂肪库中,脂肪细胞糖脂代谢基因的表达和巨噬细胞免疫应答基因的表达呈负相关关系。值得注意的是,小鼠白色脂肪细胞中对胰岛素抵抗的发生会导致巨噬细胞发生促炎反应。因此,脂肪组织炎症有可能是对脂肪细胞中已出现的代谢功能障碍的一种局部适应。脂肪细胞中不同的代谢损伤可能会产生不同的局部后果。这一观点可能有助于解释一些具有争议性的研究结果,例如在喂养高脂饮食的Cidec敲除小鼠中发现,在没有WAT炎症的情况下也可能出现胰岛素抵抗。
生物活性脂质和脂质运载蛋白分泌。脂肪细胞除了产生脂肪酸外,还可以分泌其他生物活性脂质分子来调节全身胰岛素敏感性(图7)。单不饱和脂肪酸棕榈油酸(C16:1n-7)是人类血液和脂肪组织中丰度第二高的单不饱和脂肪酸。棕榈油酸被鉴定为一种脂肪细胞特异的、DNL衍生的脂肪酸,在小鼠模型中具有胰岛素增敏特性。然而,在人体中,循环棕榈油酸水平和胰岛素敏感性之间的正相关关系仍有争议。循环棕榈油酸盐在主要来源组织(WAT和肝脏)中的合成差异,以及啮齿动物和人类之间DNL的差异,或许可以部分解释上述争议。最近一项针对非糖尿病个体的纵向研究表明,在调整了潜在干扰因素(特别是NEFA)后,循环棕榈油酸水平是胰岛素敏感性的独立决定因素。目前需要大规模干预研究来验证棕榈油酸在维持胰岛素敏感性方面的推动作用。在脂肪细胞特异性过表达GLUT4小鼠的WAT和血清中发现一类新的脂肪酸—FAHFAs(羟基脂肪酸酯)。
FAHFAs可以储存在脂肪细胞TAGs中,并被脂解动员。临床研究表明,在胰岛素抵抗个体的血清和脂肪细胞中,FAHFAs(包括棕榈酸和硬脂酸)水平较低。FAHFAs通过促进胰岛素分泌,增加脂肪细胞葡萄糖摄取,抑制WAT炎症和肝脏葡萄糖生成,对胰岛素敏感性具有有益影响。然而,在小鼠中给予FAHFAs后所导致的胰岛素敏感性改变在各研究中结论矛盾。在研究这种脂类时存在方法学上的问题,需要实验室之间的交叉验证。在WAT中,FAHFA水平由ChREBP控制。AIG1(苏氨酸水解酶)和ATRP已被证明参与FAHFAs降解,但目前尚未发现参与FAHFAs合成的酶。这两种苏氨酸水解酶的共同缺失导致WAT中(而非血浆中)FAHFA水平升高,但不能恢复高脂饮食小鼠的胰岛素敏感性。目前已经合成出一种能在体内给药的苏氨酸水解酶抑制剂,但长期给予这种抑制剂能够在胰岛素抵抗的啮齿动物模型提高胰岛素敏感性,尚待评估。在冷暴露下,棕色和米色脂肪中会产生亚油酸的一种氧化代谢物—12,13-diHOME,它通过增加脂肪细胞对脂肪酸的吸收来促进机体产热。在具有不同脂肪重量和糖耐量的人类队列中发现,血浆12,13-diHOME水平与体重指数和胰岛素抵抗呈负相关。除了对BAT的旁分泌作用外,运动也能引起12,13-diHOME分泌增加,促进骨骼肌对脂肪酸的摄取。神经酰胺是参与胰岛素抵抗的潜在脂质介质。在脂肪细胞中,抑制神经酰胺合成或促进神经酰胺降解可通过降低脂肪组织炎症和肝脏脂肪变性,促进全身代谢改善。值得注意的是,神经酰胺代谢的调节在肝脏中也起到相似作用,并且这类脂质在WAT和肝脏之间充分交换来维持代谢稳态。
脂肪细胞还分泌大量脂质运载蛋白,负责运输脂肪酸和其他脂类物质。一个典型的例子就是脂肪酸结合蛋白FABP4,它既能调控胞内脂解作用,也能在细胞受到脂解刺激时,通过非经典途径分泌。循环FABP4能激活糖异生,刺激肝脏葡萄糖生成,促进肥胖小鼠糖尿病的发展。RBP4(视黄醇结合蛋白)是另一种破坏胰岛素敏感性的脂质运载蛋白。在脂肪组织葡萄糖转运缺陷的小鼠中,RBP4表达上升。RBP4通过影响代谢和炎症促进胰岛素抵抗的发展。
WAT代谢的靶向治疗
研究发现,靶向作用于WAT代谢的药物对T2DM很有疗效,即使患者体重并未发生明显下降(图7)。噻唑烷二酮类(TZDs)类药物是一个典型例子:TZDs包括罗格列酮和吡格列酮,可作用于对脂肪分化必需的核因子PPARγ。尽管TZDs是一种高亲和力的PPARγ合成激动剂,能有效激活脂肪分化,但内源性PPARγ配体的鉴定仍然是业内古老而尚未解决的问题。根据这类核受体的配体性质,PPARγ配体被预测为是脂质或其衍生物,其中二十烷酸和脂肪酸代谢物被认为是其天然配体。TZD促进WAT中的脂质存储,改善脂肪细胞的分泌谱并减少WAT炎症,从而增加机体的胰岛素敏感性。尽管TZD因存在安全性问题而未能被广泛使用,大量证据表明TZD除了可以降低血浆葡萄糖和提高胰岛素敏感性外,还可以改善动脉粥样硬化、心血管疾病和非酒精性脂肪性肝炎。总之,这些研究表明,作用于WAT的药物具有治疗糖尿病和降低心血管代谢疾病风险的潜力。
控制WAT释放脂肪酸是改善胰岛素敏感性的有效途径。在小鼠中,使用HSL或ATGL特异性抑制剂长期抑制脂质分解可提高胰岛素敏感性。抗脂解性Gi偶联GPCR激动剂被推测具有类似作用。例如,烟酸属于该类药物,其通过HCAR2(也称为GPR109A)导致NEFA水平急剧下降(图4)。然而,烟酸治疗所介导的慢性降脂作用也通过不依赖HCAR2的机制起作用。长期烟酸治疗并没有促进血糖改善,反而破坏了血糖平衡,这可能是由于长期烟酸治疗导致的机体耐受和/或烟酸快速洗脱后NEFA水平大幅反弹所导致。通过间歇性给药方式能够成功维持烟酸降低NEFA和增强胰岛素敏感性的能力。在进食阶段定时定量给予烟酸可明显改善肥胖Zucker大鼠的新陈代谢。利用其他受体抑制脂解可能规避HCAR2激动剂带来的耐受和NEFA反弹问题。
与HCAR2类似,HCAR1(也称为GPR81)也是一种抗脂解性Gi偶联GPCR。给肥胖和胰岛素抵抗的小鼠长期服用HCAR1激动剂可增加胰岛素敏感性和抗糖尿病作用,但不改变体重。然而,HCAR1激动剂会激活肾微血管中的HCAR1,导致高血压,因此未能在人体实验中继续测试。总之,这些结果表明抑制脂解作用有助于改善胰岛素敏感性。同时,小鼠实验证明,抑制脂肪细胞的脂解作用还可有效抵抗癌症相关的恶病质发展。
相反的,激活脂解作用的同时增加脂肪酸利用是调节血糖水平和胰岛素敏感性的另一种策略。从WAT中动员的脂肪酸是促进能量消耗的关键底物。这一过程在衰老过程中可能发生了改变,由此导致能量消耗途径各组分功能衰退。近期在女性中进行的纵向研究发现,皮下WAT儿茶酚胺诱导的脂解能力随年龄增加而下降。只有在脂肪酸利用不受限的情况下,刺激脂肪细胞脂解才可能成为一种治疗手段,否则,脂肪酸很可能沉积在非脂肪组织中,从而加剧胰岛素抵抗。因此,在人类和啮齿动物中对比分析脂解激活作用时应更加严谨,因为相对于人类,小鼠和大鼠有大量活跃的BAT,具有更强的氧化脂质能力。同时,由于脂肪库β3-肾上腺素受体的分布具有种间差异,这些因素共同导致β3-肾上腺素能激动剂在啮齿动物中的有益作用远大于人类。
靶向脂肪组织还能够通过增加WAT棕色化或加速WAT中的底物循环和非UCP1依赖的生热过程,安全的增加机体能量消耗(图5)。然而,能量消耗大于产生的负平衡似乎是体重减轻的前提条件;而单独增加脂肪酸氧化对总体肥胖和体重影响并不大。此外,在成年人中,在不同脂肪组织中靶向不同产热途径对能量消耗增加的相对贡献尚未明确。近期在白色脂肪细胞中发现的其他途径也很有潜力。例如,在白色脂肪细胞中激活DNL途径与人类胰岛素敏感性之间具有强正相关性,因此被认为是有益的。然而,在脂肪肝发展过程中肝DNL会增加,所以肝脏中DNL的激活被认为有害的。由于HSL在肝脏中的表达量非常低,因此可能可以通过破坏HSL和ChREBP之间的相互作用,在脂肪细胞中特异性增强DNL和胰岛素信号通路。
从上述TZD、HCAR2和HCAR1激动剂的例子中可以看出,靶向WAT新陈代谢的分子可能因为安全性问题而无法进入临床使用。将来,开发更具针对性的药物或将药物的作用限制在特定组织或细胞类型上可能可以避免脱靶效应。具有组织特异性表达的基因靶点在已上市的非肿瘤药物中很多,但在尚处于早期临床试验的药物中却很少。因此,在考虑新的治疗靶点时,应优先考虑那些在脂肪细胞中高度或特异表达的靶点基因。
结论和未来展望
脂肪研究中的一个突出问题是WAT的异质性,这至少包含以下四个维度。首先,WAT分布在许多不同的脂肪库中,它们的解剖位置和功能各不相同。皮下和内脏脂肪库通常被认为在胰岛素抵抗和糖尿病发展中具有相反作用。但在人类中,两种脂肪细胞的全面代谢特征对比分析数据仍缺乏。在骨髓、血管周、乳腺、心外膜、关节、皮肤、眼眶后和足底等WAT较小脂肪库中的脂肪细胞对整体代谢和器官功能的影响尚未研究清楚。其次,WAT存在性别差异,并且在整个生命历程中发生着动态变化。衰老相关和性别特定的生理状态,如怀孕、哺乳和绝经,都伴随着脂肪细胞代谢的变化,而这些变化尚未得到很好的表征。考虑到这两个维度的异质性,将小鼠中的研究结果应用到人类时要充分考虑小鼠与人类的差异而极其谨慎地进行。
每个脂肪库的血管基质组分(SVF)中都包含许多不同的细胞类型,从而赋予了异质性的第三个维度(BOX1)。各个脂肪库之间免疫细胞浸润、血管和神经分布差异很大。脂肪细胞本身具有不同的颜色,即白色,棕色和米色,它们具有不同的内在特性。一个脂肪库内各个类别脂肪细胞的组成多样性构成了第四个维度。目前已发现多个白色脂肪细胞亚群具有不同代谢表型,并且在受到外源性刺激后的反应也不同。最近的研究表明,在代谢受损的肥胖个体的皮下WAT中富集了一群缺乏脂解性β2-肾上腺素能受体的人类脂肪细胞。同样,并非所有米色脂肪细胞都具有相似的代谢特征。例如,在缺乏β-肾上腺素信号通路的小鼠中鉴定出了具有高葡萄糖摄取和氧化能力的米色脂肪细胞亚群。这个亚群在人类脂肪库中是否大量存在仍不清楚。单核RNA测序技术为小鼠和人类WAT中的脂肪细胞异质性提供了新的信息。例如,通过该技术发现一群能产生乙酸的脂肪细胞亚群,会降低邻近脂肪细胞的产热能力。这种亚群在人体内比在小鼠脂肪组织中更具代表性,因此可能与人体脂肪组织的能量耗散能力较低的现象有关。
节律与代谢灵活性之间的关系越来越受到重视。除了激素、温度和摄食行为的波动外,代谢组织中正常代谢节律的建立还取决于其内源性生物钟。小鼠和人类WAT均随着昼夜节律模式表现出基因表达的巨大差异。这些节律的破坏会导致生理过程与内部时钟不一致,促进胰岛素抵抗的发生。此外,肥胖和胰岛素抵抗与代谢灵活性降低有关,也就是说,无法在饱食和禁食状态下在碳水化合物和脂质利用之间进行灵活切换。然而,长期强迫性的利用某一特定能量底物与正常生理过程相背。所以,将食物和药物摄取控制在特定时间段可能可以避免在对代谢途径的长期药物干预中观察到的有害后果。
药物治疗在所有接受治疗的患者中很少产生相同疗效。TZD就是这种情况,相当一部分T2DM患者在治疗后并未显示出胰岛素敏感性的改善。T2DM是一种高度异质性疾病:基于六个简单变量的聚类分析确定了具有不同疾病进展和糖尿病并发症风险不同的五个T2DM亚型。最近的研究表明,WAT功能对于肥胖个体的全身胰岛素抵抗和糖尿病发展很重要。表征脂肪细胞代谢变化的程度可以对代谢性疾病和癌症患者进行精准分层,从而从现有的或未来开发的药物中,筛选出对患者受益最大的药物。
白色脂肪并不仅仅是存放能量的惰性器官,而具有更多的功能。我们相信WAT是治疗心血管代谢疾病和其他存在脂肪代谢异常的疾病的有力潜在靶点。未来可开发基于脂肪细胞的新型代谢性疾病治疗策略,包括将储存能量的白色脂肪细胞转换为耗能的米色脂肪细胞,利用UCP1依赖和非依赖机制增加能量消耗,促进脂肪细胞脂质存储和氧化,以及时间依赖性的激活葡萄糖和脂质利用以恢复代谢灵活性。在使用以上方法的同时,还需要结合具有减少摄食或增加能量消耗的药物治疗,以促进机体进入分解代谢状态,充分发挥脂肪细胞疗法的潜力。
关注微信公众号代谢学人
了解更多代谢前沿资讯

转载本文请联系原作者获取授权,同时请注明本文来自徐凌燕科学网博客。
链接地址:https://blog.sciencenet.cn/blog-3483272-1309230.html
https://m.sciencenet.cn/blog-40692-1342841.html
上一篇:[转载]来自徐凌燕科学网博客:Nature综述 | 产热脂肪:超越你想象(上篇)
下一篇:[转载]来自小柯生命科学网博客:NBE新方法!靶向线粒体自噬清除大脑“垃圾”治疗阿尔兹海默症