Esposti F, Johnston J, Rosa JM, Leung KM, Lagnado L (2013) Olfactory Stimulation Selectively Modulates the OFF Pathway in the Retina of Zebrafish. Neuron 79:97-110. Dowling JE (2013) Olfaction and vision meet in the retina. Neuron 79:1-3. 在2013年7月Neuron杂志有一篇非常有意思的文章.视网膜研究元老 John E. Dowling 还为此文写了引言. 在斑马鱼 (Zebrafish) 中发现,嗅球有直接神经投射到眼睛视网膜中,而且调节了视觉.尤其是对暗信号的接受敏感性上升. 这就好比闻到美食,眼睛也发亮了,更加积极寻找食物的方位.为什么是暗信号,我想大概好吃的食物都不会是闪闪发光的.水饺包子可不发光,黄金钻石亮闪闪不过不是拿来吃的.于是眼睛就直接选择性搜索食物物体. 此文使用转基因斑马鱼,在一部分视网膜双极细胞(retinal bipolar cells)中表达GCaMP(GFP based-Calcium CaM probe). GCaMP可将细胞内钙浓度变化转换为荧光信号,从而使神经细胞活动转换为可记录的光信号,属于钙成像技术的一种.将斑马鱼幼鱼暂时固定在Agarose胶中,给予不同的味道信号,观察视网膜神经细胞活动变化. 文中所用的气味是蛋氨酸(methionine).据说类似于烤肉 爆米花 以及 炒鸡蛋的味道.给与了蛋氨酸气味后,视网膜神经细胞对暗信号反应增敏.而且此种敏感性变化可被多巴胺受体(dopamine receptor) D2R抑制剂所阻断.而其他文章从形态学实验证明嗅觉神经直接刺激多巴胺能无长突细胞(dopaminergic amacrine cell),再由多巴胺能无长突细胞(dopaminergic amacrine cell( 调制双极细胞(bipolar cell)活动. 这篇文章非常吸引我在于不同感觉器官竟然不通过大脑就有直接途径.嗅觉调节了视觉,多种感官协同合作,就是 通感 (cross modal perception, is perception that involves interactions between two or more different sensory modalities .(wiki )大脑接受了多种来源的信息. 对各种信息整合后, 大脑做出它认为最佳的判断,然后提升到意识. This is the brain's effort to provide the consciousness with its best guess about the incoming information (McGurk effect, wikipedia, ^ Jump up to: a b O’Shea, M. (2005). The Brain: A Very Short Introduction. Oxford University Press ) 曾经有理论声称所谓的情人眼里出西施也有嗅觉的功劳.当然在进化上,嗅到味道然后眼睛就更灵的鱼儿一定有生存优势.在2012年FESEB会议上另外一个实验室的poster也是关于嗅觉直接调制视觉,我印象很深刻.立意一致.而这篇文章实验要更加直观.在活体动物直接记录神经活动.也许斑马鱼通体透明,动物个体也小.实际上这篇文章所用的GCaMP2 和GCaMP3已经不是最新款的GCAMP了,2012年有GCaMP5 而今年八月Nature也发表了GCaMP6.GCaMP6的确非常灵敏,可以在小鼠视网膜皮层观察神经细胞树突棘dendritic spine上的变化,简直超乎想象.当然这篇文章中GCaMP2也许已经足够了.
博主按:本文是在我无所事事的时候写出来的。花在这些方面的精力本该用于干更有意义的事情。这篇文章与我现在的研究方向毫无关系,纯属个人兴趣爱好之作。本想将其发表于某个学术刊物,但是限于本人的水平和能力,遂放弃了这一念头。现贴出来供大家阅览,也欢迎专业人士批评指正。也许将来某一天我会从事这方面的研究工作,也许不会。 Abstract: 与大多数脊椎动物相比,昆虫更依赖嗅觉完成其生命各阶段的活动。由昆虫其个体小,神经网络结构相对简且和哺乳动物基本相同,亦是研究生物嗅觉机制的模式材料。嗅觉神经传导途径由气味分子与嗅觉受体结合开始,嗅觉受体神经元将信号传递到嗅球,信号经触角叶中局部神经元调谐后,再经投射神经元传导昆虫的蘑菇体和前脑。昆虫的嗅觉的神经传导是近年来嗅觉研究的热点之一。本文主要总结了昆虫触角叶在嗅觉信号整合和传递中的重要作用。 对于人来说,关于嗅觉有一个简单的问题:你为什么(不)喜欢这种味道?要完美地回答这个问题也许还需一定时间。研究任何感觉系统均会涉及到感受、传导、响应等一系列特征性的活动,嗅觉系统即是感受气味化合物,经嗅觉受体神经将感应信号传输至大脑,并由大脑处理后传至效应部位作出反馈的一系列的神经活动。昆虫与脊椎动物有相似的分级嗅觉传导系统。其由嗅觉受体神经元( olfactory receptor neuron, ORN )投射到触角叶中形成一个膨大的神经纤维球( glomerulus ),该神经纤维球与哺乳动物的嗅球( olfactory bulb, OB )相似。每个神经纤维球发射出一至两条投射神经元( project neuron, PN )到蘑菇体或前脑;此外,昆虫与哺乳动物感知到的外部环境大致相同,整个群体在长期的进化过程中形成了大体一致且稳定的嗅觉系统,该系统在昆虫取食、寻找配偶、产卵等一系列重大生命活动中发挥重要的作用,因此研究昆虫嗅觉系统可以阐释嗅觉发生过程中的一些普遍机理,以及嗅觉活动与整个生命活动之间的联系,为高等动物特别是人的嗅觉研究作铺垫。 在昆虫体内,刺激嗅觉器官产生嗅觉的化学分子转变成嗅觉受体神经电信号是一个复杂的生理过程。气味分子经过嗅觉感受器上的小孔进入感受器内的淋巴液内环境,与淋巴液中的气味结合蛋白结合在一起。气味结合蛋白最早被发现存在与蛾类的感觉器淋巴液中。目前发现昆虫体内主要存在两种气味结合蛋白,一种只与普通气味结合被称为普通气味结合蛋白( general odour binding protein, GOBP ),这种蛋白在多数昆虫雌雄个体中均存在。而另一种则特异性结合雌性信息素,被称为性信息素结合蛋白( pheromone binding protein, PBP )。这类蛋白一般存在于雄性个体中,但在某些少数的鳞翅目和双翅目昆虫两种性别中亦有存在。此外, GOBP 和 PBP 的分布也不同: GOBP 多数分布腔锥状感器中,而 PBP 则分布于感应信息素的毛形感器中。气味结合蛋白将气味分子携带到嗅觉受体神经末梢与气味受体结合后,激活相应的离子通道,嗅觉受体神经元立即产生动作电位,将嗅觉信息传递到下一级嗅觉中枢。在此之间,气味分子亦在很短的时间内失活。最近的研究表明,果蝇的信息素 cVA 与信息素结合蛋白 LUSH 结合,改变 LUSH 的结构 ( Laughlin, Ha et al. 2008 ) ,变构的 LUSH 与另一个跨膜蛋白 SNMP 共同作用于嗅觉受体激活胞内信号通路 ( Jin, Ha et al. 2008 ) 。值得注意的是,在此过程中相关 ORN 的动作电位并不是由信息素直接作用于嗅觉受体而引起的。然而这一配体联合机制是否可以解释其他信息素和普通气味的信号转导还需进一步研究 ( Liang and Luo 2010 ) 。 将化学信号转变为电信号的关键在于气味受体。气味受体是近年来嗅觉感受和传导的研究热点之一,目前任然存在一些争议。研究已证实哺乳动物的嗅觉受体属于 G 蛋白偶联受体( G-protein-coupled-receptor, GPCR )家族 ( Buck and Axel 1991 ) ,其具有 7 个跨膜 α- 螺旋结构,分别由三个胞外和三个胞内的环状结构相连,受体整段肽链的氨基端在胞外,羧基端在胞内;而昆虫的嗅觉受体与哺乳动物的嗅觉受体的同源性很低,约为 30% ( Nakagawa and Vosshall 2009 ) 。昆虫嗅觉受体肽链的方向和哺乳动物的相反,氨基端在胞内,而羧基端在胞外。另外与哺乳动物嗅觉受体不同的是昆虫嗅觉受体需要另一个辅助受体 ——Or83b 才能发挥出完整的化学感受能力 ( Benton, Sachse et al. 2006 ) 。 Or83b 在昆虫中普遍存在,且具有极高的同源性,它和其他的气味受体形成一个异二聚体的配体门控通道( ligand gated channel ) ( Benton, Sachse et al. 2006 ) ,然而目前有待进一步研究的是 ORN 的动作电位是由气味受体直接作用于配体门控通道引起的离子流还是由 cAMP 第二信使途径引起的。 1 昆虫的触角叶的构成 当嗅觉信号经过 ORN 传递至昆虫的第一级嗅觉中枢 —— 触角叶( Antennal lobe , AL )时,信号传递便开始受到神经调控。近十年来,随着研究手段和方法的不断改进和创新,嗅觉信号的神经传导和调节的相关研究已经突飞猛进发展。研究对象主要是双翅目、膜翅目、直翅目和鳞翅目的昆虫。 昆虫触角叶与脊椎动物的嗅球一样是嗅觉信号处理的一级中枢。由无数个嗅觉受体神经纤维球神经纤维球组成,神经纤维球是嗅觉信息处理的的基本功能单位。嗅觉神经纤维球在昆虫体内的数量和昆虫的进化程度或者是生存环境有关。果蝇每个触角叶中大有有 50 个神经纤维球,蜜蜂中大约有 160 个,而蝗虫则有多大 1000 的微型神经纤维球 ( Wilson and Mainen 2006 ) 。多数情况下,昆虫雌雄两性的触角叶没有差异,而在某些蜚蠊目和鳞翅目的昆虫中触角叶存在雌雄二型性,如烟草天蛾的雄性中存在一个扩大型神经纤维球复合物( macroglumerular complex, MGC )。研究证实 MGC 与感受雌性性信息素有关 ( Rospars and Hildebrand 1992 ) 。 每个神经纤维球有 1-2 条输出神经 —— 称为投射神经元( project neuron, PN ),将汇总并经过一定处理的信号传输至昆虫的蘑菇体和前脑。神经纤维球之间由无数的无轴突的局部神经元( local neuron, LN )连接, LN 和其他的 LN 、 ORN 、 PN 之间均有连接。 每个神经纤维球由表达相同嗅觉受体的一组嗅觉受体神经元扩大形成。在昆虫中,同一组感受器中神经元可能不表达一种 OR ,但是在 AL 中汇聚在一起形成膨大神经纤维球的 ORN 则表达同一种受体。在单条神经元中一般情况下至表达一种受体,但是有例外;但是在哺乳动物中一条嗅觉神经元只表达一种受体。嗅觉神经元的数量可能与动物的进化程度有关,在小鼠中约有 200 万 ORNs ,果蝇成虫中大约有 1300 个 ORN ,在蝗虫中每个 AL 接受来自于约 90000 条 ORNs 输入。不同发育阶段和生活环境下的同一种动物 ORNs 也不同。果蝇幼虫只有 21 条 ORN ,可能与其整个幼虫期生活在腐烂水果中有关,相对狭小的生活环境可能使幼虫期的果蝇进化出简单的嗅觉神经系统。而成虫期的生活环境相对复杂,个体更依赖嗅觉辨别复杂的环境,因此而进化相对复杂的嗅觉系统。但是也有研究称果蝇嗅觉系统中存在冗余 ( Fishilevich, Domingos et al. 2005 ) ,实验证实,单一的 ORN 即可使果蝇幼虫感受化学物质并作出相应的行为反应,事实上同一化合物可以同时结合多种嗅觉受体并引起相应的 ORN 产生动作电位,但是这可能有助于探测较低浓度的气味物质。这样的汇聚作用可能有助于提高信噪比(见后) ( Wilson and Mainen 2006 ) 。 投射神经元( project neuron, PN )作为输出神经元将初步处理后的信号传递至昆虫下一集嗅觉中枢蘑菇体和前脑中。在鳞翅目,双翅目,膜翅目昆虫中,通常情况下一条 PN 支配一个神经纤维球,这些 PN 也是研究时电生理记录的主要对象。但偶尔也会出现一条 PN 支配两个神经纤维球的情况。例外的是在蝗虫中,所有的 PN 均支配多个神经纤维球。 ORN 和 PN 均以乙酰胆碱作为神经递质,表明 ORN 和 PN 传导兴奋性的神经电信号。 在 AL 中 ORN 和 PN 分别作为输入和输出神经元,那么在信号调节中起关键作用的则是局部神经元( local neuron, LN )。 LN 的显著特征是没有轴突,广泛分布于触角叶中,调节 ORN 输入的嗅觉信号。以前认为 LN 只连接相邻的两个神经纤维球,随后证明相距较远的神经纤维球也有 LN 连接,单条 LN 可扩展至整个 AL 。然而解剖和生理实验表明,某些 LN 只支配特定的神经纤维球。且多数 LN 释放 GABA 作为神经递质,表明 LN 参与抑制性的神经调节,被称为抑制性局部神经元( iLN )。 LN 的抑制效应首次在烟草天蛾中得到证实,并观察到 ORN 直接向 LN 输入信号 ( Christensen, Waldrop et al. 1993 ) 。然而最近两实验室几乎同时发现, AL 亦存在兴奋性的 LN ( eLN ) ( Olsen, Bhandawat et al. 2007 ; Shang, Claridge-Chang et al. 2007 ) 。 LNs 的神经递质种类、链接方式、以及生理特性均有很强的多样性;尽管多数 LN 是 GABA 能的,但是少数还是采用其他物质为递质,如乙酰胆碱、多巴胺、谷氨酸等,或者同时释放多种递质。 2 触角叶提高信噪比和增益控制的机制 动物分级嗅觉系统的优势在于能对信号进行适当分散或汇聚,假如化学感受物质和 ORN 是一比一的关系。那么则会存在一下情况:第一,当感受某种物质的 ORN 受损伤或者发生可塑性改变后,动物就失去了感受某种物质的能力;第二,过多的平行神经元会造成神经元数量增加,扩大脑容量,且使信号处理更困难,动物进化上处于劣势。然而自然界中化学感受物质除了物理化学性质上的差异各种物质的浓度也有差异,昆虫的嗅觉系统如何适应这种差异呢? 信噪比( signal-to-noise ratio )和增益控制( gain control )是保证嗅觉信号准确快速传递到下一嗅觉中枢的重要神经活动。前者重在放大微弱的信号;后者重在抑制较强的信号。在脊椎动物和昆虫中,一个典型的提高信噪比的内在嗅觉结构基础便是 ORN 的汇聚作用。多条表达相同 OR 的 ORN 汇聚成一个膨大的神经纤维球,而一个神经纤维球通常只发出 1-2 条 PN 至嗅觉更高级中枢 ( Hallem, Ho et al. 2004 ) ,进一步研究表明在 PN 中传递的来自 ORN 的兴奋性突触后电流 (excitatory postsynaptic currents, EPSCs) 包括两部分,一部分由突触前 ORN 递质释放引起,另一部分则是来自于与 ORN 连接的 eLN ( Kazama and Wilson 2008 ) 。此外电生理实验还表明 ORN 和 PN 之间的信号传递呈非线性关系,即使当 ORN 受到微弱的刺激其信号峰靠近基线几乎检测不到时,在 PN 中却能检测到较为强烈的信号 ( Bhandawat, Olsen et al. 2007 ) 。 与此同时,神经系统往往还面临频繁的、相同的信号刺激,那么这些“多余的”信号是怎么被屏蔽掉的呢? Bhandawat et al . (2007) ( Bhandawat, Olsen et al. 2007 ) 同时记录 ORN 和 PN 的电信号发现当高浓度的化感物质刺激 ORN 产生强烈的信号时,在 PN 中检测到的信号要比最最初的刺激弱得多,且 PN 主要传递 ORN 动作电位上升阶段的信号,即在信号传递的同时 PN 比 ORN 更先到达峰值。增益控制的另一机制在于 ORN 和 PN 之间的短期抑制( short-term depression )。随着 ORN 放电频率增大,突触后电流却越小,原因可能是过快的脉冲导致突触前递质囊泡释放减少 ( Kazama and Wilson 2008 ) 。此外, eLN 在增益控制方面亦发挥了重要作用。 ORN 上的 GABA A 和 GABA B 受体共同参与调节。 GABA A 主要负责信号起始后的短暂抑制,而 GABA B 主要负责信号传递后期的较长一段时间的抑制作用 ( Wilson 2005 ; Olsen and Wilson 2008 ) 。最近的研究表明, LN 的侧抑制调节增益控制主要有以下三特征: 1, 靶标部位至少在 ORN 末端 ( Olsen and Wilson 2008 ) ; 2, 抑制强度与总的 ORN 输入相 ( Root, Masuyama et al. 2008 ) ; 3, 不同神经纤维球抑制强度不同 ( Asahina, Louis et al. 2009 ) 。但是 AL 中的 LN 网络是如何精细地调节为数众多的神经纤维球,并准确控制其向下一级神经中枢输入的还需进一步研究。 提高信噪比以及增益控制即是嗅觉神经系统将微弱的信号和过强的信号控制在一个同等的数量级的机制,这可能有利于下一级嗅觉中枢对嗅觉信号的进一步处理。同时这也可能保证了信号传导的真实性和客观性。如微弱的信号被放大,以至于与该信号相关的信息不被丢失。然而关于嗅觉系统信息传导的信号保真度还没有相关报道。从宏观的角度看,昆虫所处环境中的气味感受物质随时间和空间不断变化。这一嗅觉机制使得昆虫能够迅速地感受并调节生理及行为活动以适应变化。 3 触角叶中嗅觉信号传导和整合的差异 3.1 不同发育期的嗅觉系统差异 虽然在昆虫生活史中不同发育时期参与嗅觉感受与记忆的器官——触角叶、蘑菇体、前脑等均存在于幼虫和成虫中,但是参与构成这些器官的内部神经网络却不同。以果蝇为例,在果蝇幼虫中仅存在约 21 个嗅觉受体神经元,触角叶约 21 个神经纤维球,约有 21 条投射神经元;各级嗅觉神经数量几乎呈现一比一的关系,而成虫则有约 1300 条嗅觉受体神经元,汇聚成约 43 个膨大的神经纤维球,这些神经纤维球又发出约 150 条投射神经元到下一级嗅觉中枢 ( Ramaekers, Magnenat et al. 2005 ) 。这表明成虫的嗅觉处理存在一个将信号收集并放大的一个过程,抑或是成虫嗅觉系统存在冗余 ( Ramaekers, Magnenat et al. 2005 ) 。此外这种差异可能与生活环境与功能差异有关。果蝇幼虫主要以取食腐败的水果,其整个幼虫期的生活范围可能仅仅局限于某个腐败的水果内,其主要任务便是完成化蛹。因此整个过程可能不需要一个复杂的嗅觉系统。然而成虫期的生活范围要比幼虫期更广且要完成一些生活史中关键的任务——寻早配偶、选择合适的产卵地等。而这一系列过程均需要经过对自然界中有关的化学物质进行“搜集”和“评估”。因此需要一个较为复杂的嗅觉系统。 3.2 基于不同感受功能的嗅觉神经传导差异 在蛾类昆虫中,信息传导的化学信号转化为神经电信号之前就已经出现了差异。即如前面所述,信息素和和普通气味分别与信息素结合蛋白( PBP )和普通气味结合蛋白( GOPB )结合,并转运至相应的嗅觉受体上。那么我们不禁会问嗅觉传导的下一步过程中是否也会存在着(甚至更大的)差异呢?是否意味着嗅觉信息传导因适应不同的生理需求(如繁殖后代、寻找食物充饥等)而存在差异呢? 在雄性烟草天蛾 AL 中存在一个特殊的神经纤维球,即扩大性神经纤维复合体( MGC )。它由一组感受雌性信息素的 ORN 汇聚形成。但是 MGC 在雌性个体中并不存在。表明 MGC 可能与特异性信息素感受有关。进一步的钙影像和电生理实验表明,有关信息素的信号只经过 MGC 传递到下一级嗅觉中枢,而普通气味则经过一般的神经纤维球传递 ( Christensen and Hildebrand 1987 ; Anton and Hansson 1994 ) 。然而行为学实验证明,植物气味物质能加强雄蛾对信息素的感受 ( Ochieng, Park et al. 2002 ; Reddy and Guerrero 2004 ) ;另一方面,雌蛾也倾向于在偏好的绿色植物上求偶,以增加成功率 ( Christensen and Hildebrand 2002 ) 。那么蛾类嗅觉系统对普通气味和信息素的编码是纯平行的呢?还是相互关联的呢? Christensen ( 2000 ) ( Christensen, Pawlowski et al. 2000 ) 等用微电极证实了普通气味和信息素编码存在联系,然而要弄清其机理还需进一步研究。此外,相似的嗅觉系统还存在于蚂 蚁、蜜蜂、蟑螂等昆虫中。 在果蝇中雌雄和雄性 AL 中的神经纤维球在数量上没有差异。但是在雄性个体中的 DA1 和 VA1lm 两个神经纤维球要比雌性个体中的稍大 ( Kondoh, Kaneshiro et al. 2003 ) 。与蛾类昆虫不同的是果蝇不使用信息素进行长距离交流,且它们只使用一个短距离交流的信息素—— cis-vaccenyl acetate 或者( -11-octadecenyl acetate ) ( Amrein 2004 ) 。该信息素在两性个体中受同一嗅觉受体 Or67d 编码,对雌性成虫表现出吸引作用,对雄性成虫表现出趋避作用 ( Kurtovic, Widmer et al. 2007 ) 。然而在雌雄个体的 AL 结构基本相同的情况下,这种对立的行为差异是如何实现的?有何更深层次的神经结构差异呢? Datta ( 2008 )等利用双光子激光扫描显微镜( two-photon laser scanning microscopy )结合光激活绿色荧光蛋白( photoactivatable green fluorescent protein )标记发现由 DA1 输出的 PN 分别投射到雌雄个体前脑的不同区域,这一解剖学上的差异造成了雌雄果蝇对其信息素的不同反应 ( Datta, Vasconcelos et al. 2008 ) 。 果蝇化学感受另一特殊之处在对 CO 2 的感受。该过程与其他气味感受相比表现得十分独立且简单,不需要联合编码。参与 CO2 的是一对特殊的受体—— (Gr21a/Gr63a) ( Jones, Cayirlioglu et al. 2007 ; Kwon, Dahanukar et al. 2007 ) 。其神经元共同支配 AL 中的一个神经纤维球—— V ( Suh, Wong et al. 2004 ) ,且不表达 GABA 受体,这表明由 ORN 向 PN 传输的 CO 2 信号几乎不受到抑制性的 LN 调控 ( Root, Masuyama et al. 2008 ) ,直接将上一级嗅觉信号传递至下级嗅觉中枢。 CO 2 是引起果蝇应激反应的化学感受物质 ( Suh, Wong et al. 2004 ) 。受到干扰的果蝇会产生较高浓度的 CO 2 ,对其他果蝇具有趋避作用。因此进化出简单的快速的 CO 2 感受系统有助于果蝇作出快速应急反应,提高其在自然界的生存能力。 4 触角叶中传导系统的可塑性与嗅觉感受 关于哺乳动物嗅觉系统的可塑性已有不少研究 ( Wilson, Best et al. 2004 ; Tyler, Petzold et al. 2007 ; Wilson 2010 ) 。长期以来人们认为突触的可塑性是神经系统储存信息的机制。在嗅觉系统中,短期的突触可塑性存在于嗅球和 梨状皮质中,这种可塑性可以储存短时的嗅觉信息和响应重复的信号输入,并能在复杂的气味中精细调节嗅觉活动 ( Wilson, Best et al. 2004 ) 。然而昆虫的嗅觉可塑性的神经机制的相关研究则相对较少。一项研究表明,果蝇的嗅觉系统中的嗅觉神经元和投射神经元很稳定 ( Berdnik, Chihara et al. 2006 ) 。研究人员无论是使用内源表达的毒素失活嗅觉感受神经元还是物理切除触角,其相应的和其他的投射神经元都很稳定,也不会改变投射神经元在高级神经中枢的分布。反之失活投射神经元,嗅觉感受神经元亦很稳定。 事实表明构成昆虫嗅觉传导主要框架的嗅觉感受神经元和投射神经元很稳定,那么什么结构使昆虫嗅觉具有可塑性呢?如提前暴露在信息素中的 Spodoptera littoralis 雄虫较未接触信息素的对照雄虫对信息素更敏感 ( Anderson, Sadek et al. 2003 ; Anderson, Hansson et al. 2007 ) ;被暴露在其近缘种 Heliothis virescens 雌虫信息素中的 Heliothis subflexa 雌虫信息素主要成分含量会发生改变 ( Groot, Classen et al. 2010 ) ;未接触过信息素中的雄性果蝇比长期暴露在信息素中的有更强的好斗性 ( Sachse, Rueckert et al. 2007 ; Liu, Liang et al. 2011 ) 。 Sachse, Rueckert et al. 2007 等发现果蝇长期暴露于较高浓度的 CO2 中,神经纤维球 V 的体积会增大,与此同时从 V 神经纤维球输出的投射神经元向侧角的输入减少。这种神经纤维球体积的变化在特定的时期是可逆的,当 CO2 的浓度降低后,神经纤维球 V 的体积则会变小。这种体积变化还可以推广到其他的神经纤维球。然而这种体积变化后隐藏着什么深层次的机理呢? 触角叶内的神经纤维球之间有丰富的局部神经元连接,这些神经元由兴奋型和抑制型两类构成 ( Olsen, Bhandawat et al. 2007 ; Shang, Claridge-Chang et al. 2007 ) 。它们在嗅觉的精细调节和嗅觉可塑性中起重要作用。兴奋型神经元、抑制型神经元、嗅觉感受神经元和投射神经元四者之间相互联系,在触角叶中构成一个小型的神经网络 ( Wilson 2011 ) 。但是通常认为嗅觉感受神经元中的直接传递给与其同一神经纤维球的投射神经元 ( Root, Semmelhack et al. 2007 ) 。快速切除嗅觉感受神经元后立刻记录其相应的上一级投射神经元对气味的反应,发现气味引起的投射神经元去极化程度很低。而切除 2 天后记录投射神经元去极化程度则很高。进一步研究发现,在神经胶质细胞的介导下,切除嗅觉感受神经元增强局部神经元突触之间联系 ( Kazama, Yaksi et al. 2011 ) 。然而在嗅觉感受神经元和投射神经元都完整的情况下,局部神经元是否会随着动物的经历不同而发生相应的改变还需进一步研究。 5 结论与展望 昆虫嗅觉的研究之所以得到广泛的关注,一是由于昆虫很适合用作嗅觉系统功能和发育的研究材料,可以为深入研究高等动物特别是人类的嗅觉打下坚实基础;二是由于许多农业害虫和卫生害虫的经济和社会重要性,而这些害虫中大多数是通过嗅觉信息找到寄主植物或人类的,研究昆虫嗅觉感受的分子基础,将为害虫控制提供新的途径。然而目前研究嗅觉神经传导通路还没有大规模地检测众多神经通路的活动情况,通常的做法是刺激或失活某个神经元然后再研究其临近神经元的活动。但是单个神经元是整个神经通路的一部分,其活动变化可能会影响到整个神经网络的活动。因此要彻底弄清楚嗅觉神经网络,应在其解剖学的基础上应用计算神经生物学和复杂网络科学的方法。 Amrein, H. (2004). Pheromone perception and behavior in Drosophila. Current Opinion in Neurobiology. 14 (4): 435-442. Anderson, P., B. S. Hansson, et al. (2007). Increased behavioral and neuronal sensitivity to sex pheromone after brief odor experience in a moth. Chem Senses. 32 (5): 483-491. Anderson, P., M. M. Sadek, et al. (2003). Pre-exposure modulates attraction to sex pheromone in a moth. Chem Senses. 28 (4): 285-291. Anton, S. and B. S. Hansson (1994). Central processing of sex pheromone, host odour, and oviposition deterrent information by interneurons in the antennal lobe of female Spodoptera littoralis (Lepidoptera: Noctuidae). J Comp Neurol. 350 (2): 199-214. Asahina, K., M. Louis, et al. (2009). A circuit supporting concentration-invariant odor perception in Drosophila. J Biol. 8 (1): 9. Benton, R., S. Sachse, et al. (2006). Atypical membrane topology and heteromeric function of Drosophila odorant receptors in vivo. PLoS Biol. 4 (2): e20. Berdnik, D., T. Chihara, et al. (2006). Wiring stability of the adult Drosophila olfactory circuit after lesion. J Neurosci. 26 (13): 3367-3376. Bhandawat, V., S. R. Olsen, et al. (2007). Sensory processing in the Drosophila antennal lobe increases reliability and separability of ensemble odor representations. Nat Neurosci. 10 (11): 1474-1482. Buck, L. and R. Axel (1991). A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell. 65 (1): 175-187 Christensen, T. A. and J. G. Hildebrand (1987). Male-specific, sex pheromone-selective projection neurons in the antennal lobes of the moth Manduca sexta. J Comp Physiol A. 160 (5): 553-569. Christensen, T. A. and J. G. Hildebrand (2002). Pheromonal and host-odor processing in the insect antennal lobe: how different? Curr Opin Neurobiol. 12 (4): 393-399. Christensen, T. A., V. M. Pawlowski, et al. (2000). Multi-unit recordings reveal context-dependent modulation of synchrony in odor-specific neural ensembles. Nat Neurosci. 3 (9): 927-931. Christensen, T. A., B. R. Waldrop, et al. (1993). Local interneurons and information processing in the olfactory glomeruli of the moth Manduca sexta. J Comp Physiol A. 173 (4): 385-399. Datta, S. R., M. L. Vasconcelos, et al. (2008). The Drosophila pheromone cVA activates a sexually dimorphic neural circuit. Nature. 452 (7186): 473-477. Fishilevich, E., A. I. Domingos, et al. (2005). Chemotaxis Behavior Mediated by Single Larval Olfactory Neurons in Drosophila. Current Biology. 15 (23): 2086-2096. Groot, A. T., A. Classen, et al. (2010). Phenotypic plasticity in sexual communication signal of a noctuid moth. J Evol Biol. 23 (12): 2731-2738. Hallem, E. A., M. G. Ho, et al. (2004). The molecular basis of odor coding in the Drosophila antenna. Cell. 117 (7): 965-979. Jin, X., T. S. Ha, et al. (2008). SNMP is a signaling component required for pheromone sensitivity in Drosophila. Proc Natl Acad Sci U S A. 105 (31): 10996-11001. Jones, W. D., P. Cayirlioglu, et al. (2007). Two chemosensory receptors together mediate carbon dioxide detection in Drosophila. Nature. 445 (7123): 86-90. Kazama, H. and R. I. Wilson (2008). Homeostatic Matching and Nonlinear Amplification at Identified Central Synapses. Neuron. 58 (3): 401-413. Kazama, H., E. Yaksi, et al. (2011). Cell Death Triggers Olfactory Circuit Plasticity via Glial Signaling in Drosophila. Journal of Neuroscience. 31 (21): 7619-7630. Kondoh, Y., K. Y. Kaneshiro, et al. (2003). Evolution of sexual dimorphism in the olfactory brain of Hawaiian Drosophila. Proc Biol Sci. 270 (1519): 1005-1013. Kurtovic, A., A. Widmer, et al. (2007). A single class of olfactory neurons mediates behavioural responses to a Drosophila sex pheromone. Nature. 446 (7135): 542-546. Kwon, J. Y., A. Dahanukar, et al. (2007). The molecular basis of CO2 reception in Drosophila. Proc Natl Acad Sci U S A. 104 (9): 3574-3578. Laughlin, J. D., T. S. Ha, et al. (2008). Activation of pheromone-sensitive neurons is mediated by conformational activation of pheromone-binding protein. Cell. 133 (7): 1255-1265. Liang, L. and L. Luo (2010). The olfactory circuit of the fruit fly Drosophila melanogaster. Sci China Life Sci. 53 (4): 472-484. Liu, W., X. Liang, et al. (2011). Social regulation of aggression by pheromonal activation of Or65a olfactory neurons in Drosophila. Nat Neurosci. 14 (7): 896-902. Nakagawa, T. and L. B. Vosshall (2009). Controversy and consensus: noncanonical signaling mechanisms in the insect olfactory system. Curr Opin Neurobiol. 19 (3): 284-292. Ochieng, S. A., K. C. Park, et al. (2002). Host plant volatiles synergize responses of sex pheromone-specific olfactory receptor neurons in male Helicoverpa zea. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 188 (4): 325-333. Olsen, S. R., V. Bhandawat, et al. (2007). Excitatory Interactions between Olfactory Processing Channels in the Drosophila Antennal Lobe. Neuron. 54 (1): 89-103. Olsen, S. R. and R. I. Wilson (2008). Lateral presynaptic inhibition mediates gain control in an olfactory circuit. Nature. 452 (7190): 956-960. Ramaekers, A., E. Magnenat, et al. (2005). Glomerular Maps without Cellular Redundancy at Successive Levels of the Drosophila Larval Olfactory Circuit. Current Biology. 15 (11): 982-992. Reddy, G. V. and A. Guerrero (2004). Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci. 9 (5): 253-261. Root, C. M., K. Masuyama, et al. (2008). A Presynaptic Gain Control Mechanism Fine-Tunes Olfactory Behavior. Neuron. 59 (2): 311-321. Root, C. M., J. L. Semmelhack, et al. (2007). Propagation of olfactory information in Drosophila. Proceedings of the National Academy of Sciences. 104 (28): 11826-11831. Rospars, J. P. and J. G. Hildebrand (1992). Anatomical identification of glomeruli in the antennal lobes of the male sphinx moth Manduca sexta. Cell Tissue Res. 270 (2): 205-227. Sachse, S., E. Rueckert, et al. (2007). Activity-dependent plasticity in an olfactory circuit. Neuron. 56 (5): 838-850. Shang, Y., A. Claridge-Chang, et al. (2007). Excitatory Local Circuits and Their Implications for Olfactory Processing in the Fly Antennal Lobe. Cell. 128 (3): 601-612. Suh, G. S., A. M. Wong, et al. (2004). A single population of olfactory sensory neurons mediates an innate avoidance behaviour in Drosophila. Nature. 431 (7010): 854-859. Tyler, W. J., G. C. Petzold, et al. (2007). Experience-dependent modification of primary sensory synapses in the mammalian olfactory bulb. J Neurosci. 27 (35): 9427-9438. Wilson, D. A. (2010). Single-unit activity in piriform cortex during slow-wave state is shaped by recent odor experience. J Neurosci. 30 (5): 1760-1765. Wilson, D. A., A. R. Best, et al. (2004). Plasticity in the olfactory system: lessons for the neurobiology of memory. Neuroscientist. 10 (6): 513-524. Wilson, R. I. (2005). Role of GABAergic Inhibition in Shaping Odor-Evoked Spatiotemporal Patterns in the Drosophila Antennal Lobe. Journal of Neuroscience. 25 (40): 9069-9079. Wilson, R. I. (2011). Understanding the functional consequences of synaptic specialization: insight from the Drosophila antennal lobe. Curr Opin Neurobiol. 21 (2): 254-260. Wilson, R. I. and Z. F. Mainen (2006). Early events in olfactory processing. Annu Rev Neurosci. 29 : 163-201.
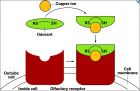
据美国 Proc. Natl. Acad. USA, DOI: 10.1073/pnas.1111297109 )的最新报道,科学家的小白鼠动物实验结果研究发现,某些嗅觉受体,需要Cu 2+ 的帮助,可以对一些特别臭的硫醇以及相关气体化合物更加敏感。这种发现可能对于没有建立嗅觉感知新模式、修改已有观念有所帮助。更为详细报道可以参看 February 8, 2012 | Latest News Metals’ Role In Odor Sensing Ne uroscience: Some olfaction receptors need the help of copper, research shows
The mathematics of taste By using ‘genetic programming’ to crossbreed algorithms, researchers help flavor companies figure out what their customers like. The design of aromas — the flavors of packaged food and drink and the scents of cleaning products, toiletries and other household items — is a multibillion-dollar business. The big flavor companies spend tens of millions of dollars every year on research and development, including a lot of consumer testing. But making sense of taste-test results is difficult. Subjects’ preferences can vary so widely that no clear consensus may emerge. Collecting enough data about each subject would allow flavor companies to filter out some of the inconsistencies, but after about 40 flavor samples, subjects tend to suffer “smell fatigue,” and their discriminations become unreliable. So companies are stuck making decisions on the basis of too little data, much of it contradictory. One of the biggest flavor companies in the world has turned to researchers in MIT’s Computer Science and Artificial Intelligence Laboratory (CSAIL) for help. To analyze taste-test results, the CSAIL researchers are using genetic programming, in which mathematical models compete with each other to fit the available data and then cross-pollinate to produce models that are more accurate still. The Swiss flavor company Givaudan asked CSAIL principal research scientist Una-May O’Reilly, postdoc Kalyan Veeramachaneni and the University of Antwerp’s Ekaterina Vladislavleva to help interpret the results of tests in which 69 subjects evaluated 36 different combinations of seven basic flavors, assigning each a score according to its olfactory appeal. For each subject, O’Reilly and her colleagues randomly generate mathematical functions that predict scores according to the concentrations of different flavors. Each function is assessed according to two criteria: accuracy and simplicity. A function that, for example, predicts a subject’s preferences fairly accurately using a single factor — say, concentration of butter — could prove more useful than one that yields a slightly more accurate prediction but requires a complicated mathematical manipulation of all seven variables. After all the functions have been assessed, those that provide poor predictions are winnowed out. Elements of the survivors are randomly recombined to produce a new generation of functions; those are then evaluated for accuracy and simplicity. The whole process is repeated about 30 times, until it converges on a set of functions that accord well with the preferences of a single subject. Because O’Reilly and her colleagues’ method produces profiles of individual test subjects’ tastes, it can sort them into distinct groups. It could be, for instance, that test subjects tend to have strong preferences for either cinnamon or nutmeg but not both. By marketing one product to cinnamon lovers and another to nutmeg lovers, a company could do much better than by marketing one product to both. “For every one of these 36 flavors, someone hated it and someone liked it,” O’Reilly says. “If you try to identify a flavor that the whole panel likes, you end up settling for a little bit less.” O’Reilly and her colleagues haven’t had an opportunity to empirically determine whether their models correctly predict subjects’ responses to new flavors. So to try to establish their model’s accuracy, they instead built another model. First, they developed a set of mathematical functions that represent subjects’ true taste preferences. Then they showed that, given the limitations of particular test designs, their algorithms could still divine those preferences. Although they developed the model purely to validate their approach, O’Reilly says, flavor researchers were intrigued by the possibility of using it to develop more accurate and efficient test protocols. “People have been playing with these techniques for decades,” says Lee Spector, a professor of computer science at Hampshire College and editor-in-chief of the journal Genetic Programming and Evolvable Machines , where the MIT researchers’ latest paper appears. “One of the reasons that they haven’t made a big splash until recently is that people haven’t really figured out, I think, where they can pay off big.” Taste preference, Spector says, “is a pretty brilliant area in which to apply the evolutionary methods — and it looks as though they’re working, also, so that’s exciting.” http://web.mit.edu/newsoffice/2012/what-smells-good-0124.html
昆虫嗅觉研究争议(一):化学感受物质是否直接与嗅觉受体结合? 昆虫的嗅觉感受和哺乳动物嗅觉感受有所不同。昆虫的化学感受主要部位位于头部类似于天线状的触角,但是不同种类的昆虫触角的形态不同,此外一些昆虫(如果蝇)的翅外缘,足末端也存在化学感受位点。 昆虫的触角上有许多纤毛状或者棒状或者其它类型的感受器。这些感受器不仅负责化学感受,部分还负责温度、湿度的感受。化学感受器的特征是表面分布有数量不一的小孔,供气味物质通过进入感受器内。化学感受器内部是淋巴液围着的嗅觉神经元,淋巴液中还存在与气味物质结合的蛋白,被称为气味结合蛋白,目前气味结合蛋白大致分为两类一类是普通气味结合蛋白,而另一种则特异性结合雌性信息素,被称为性信息素结合蛋白。 气味物质如蛾类昆虫的信息素多为长链的疏水物质,不能穿过淋巴液与受体结合,便先与气味结合蛋白结合,然后再由气味结合蛋白转运至嗅觉受体,使嗅觉受体神经元产生动作电位。 然而问题是这个动作电位是由气味物质与嗅觉受体作用产生的呢,还是由气味结合蛋白与嗅觉受体结合产生的呢? 德克萨斯大学西南医学中心的 Dean Smith 发现果蝇的信息素 cVA 与信息素结合蛋白 LUSH 结合后改变 LUSH 的结构,变构的 LUSH 和一个膜蛋白 SNMP 共同作用激发了嗅觉神经元的动作电位。该研究发现, LUSH 蛋白上有两个位点决定整个蛋白质的功能,即第 121 号苯丙氨酸和 118 号天冬氨酸,其中 121 号氨基酸的大小决定嗅觉神经元动作电位高低。 118 号天冬氨酸与 87 号赖氨酸形成盐桥,使 LUSH 失活,而当盐桥被打断后 LUSH 的构象和与 cVA 结合时的 LUSH 并无两样,因此 cVA 可能发挥了打断 118 号氨基酸与 87 号氨基酸之间的盐桥的作用。 然而这一实例是普遍现象呢,还是特殊现象?。昆虫在感受普通气味时是否着经过这种途径呢?还是是气味物质直接与受体结合?此外还需要其他膜蛋白参与么? 要解决这些问题首先要找到目标基因,然后敲除恢复验证,纯化蛋白,解析结构,找到作用位点。显然这些东西不是我们玩的。 参考文献 Jin, X., T. S. Ha, et al. (2008). "SNMP is a signaling component required for pheromone sensitivity in Drosophila." Proc Natl Acad Sci U S A 105 (31): 10996-11001. Kruse, S. W., R. Zhao, et al. (2003). "Structure of a specific alcohol-binding site defined by the odorant binding protein LUSH from Drosophila melanogaster." Nat Struct Biol 10 (9): 694-700. Laughlin, J. D., T. S. Ha, et al. (2008). "Activation of pheromone-sensitive neurons is mediated by conformational activation of pheromone-binding protein." Cell 133 (7): 1255-1265. Liang, L. and L. Luo (2010). "The olfactory circuit of the fruit fly Drosophila melanogaster." Sci China Life Sci 53 (4): 472-484.
 标签: 嗅觉
标签: 嗅觉