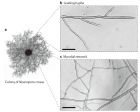
菌丝是如何生长的?压力促使菌丝生长的生物物理学研究 (美文推荐) 在一个菌落中,菌落前端的菌丝以寻找食物为目的向前生长,菌落中间的菌丝则结成三维网状结构,最大化地从菌丝周围的培养基上吸收营养物质供给生长(上图所示)。菌落的生长速度可以是很快的(大约 10-100um/min ,这要依赖于物种类型,可利用的营养物质,以及温度条件等),菌落的生长是一个涉及为使菌丝细胞快速扩张的所有细胞组分持续合成的过程。驱使真菌菌丝生长的主要作用力是——压力。 压力是一个热力学特性,影响到各种生物的生命过程。对于缺乏细胞壁的生物来说,压力可能是致命的,将导致细胞的裂解和死亡。而对有细胞壁的生物来说(大多数的细菌,藻类,真菌以及植物),内部静水压力(膨胀压)对生物的一些独立结构提供支撑作用,以及对细胞扩张、物质的吸收和其他一些过程提供一种驱使力。一个特别的例子是,在一些子囊菌和接合菌门的真菌中,孢子释放过程可以产生巨大的张力 100000 × g ( g 为地球表面的重力加速度),对于一些寄生真菌来说,这样的膨胀压力足以使其穿过寄主组织。膨胀压是由于细胞内渗透物的浓度比细胞外高而产生的渗透作用所致。细胞的生长(扩张或分化)以及细胞壁的重塑过程需要增加细胞的总量。我们所知道的大多数细胞扩张经历的机制被认为是菌丝细胞顶端延伸管延伸的尖端生长机制所致。关于尖端生长的模式系统的例子有很多,如藻类,真菌,植物的花粉管和根尖的生长。菌丝尖端的生长模式是一个梯度生长的,菌丝的尖端扩展最快,紧挨着菌丝尖端的那些就开始减慢了。 1892 年 Reinhardt 就对这一些进行过认真的研究。这样的扩张模式(尖端生长)与膨压驱动的增长是一致的。 这篇讲解菌丝到底是如何生长以及相关机制的研究的文章发表在 2011 年六月的 Nature Reviews Microbiology ,附原文,喜欢的随便下载。 How does a hypha grow? The biophysics of pressurized growth in fungi nrmicro2011.pdf
 标签: 菌丝
标签: 菌丝