单细胞,单分子测序给癌症检测带来了突飞猛进的发展。 最近nature methods发表了一篇牛逼文章,可以同时 检测基因组和转录组两个层次得变化 In those cells where chromosomal gains or losses (either reciprocal or nonreciprocal) were seen at the genomic level, we observed concomitant increases and decreases in chromosome-wide relative gene expression levels after GT-seq analysis, which established for the first time (to our knowledge) that the effects of gene expression dosage can be rapidly established after the acquisition of aneuploidies during a single cell division. 下面给生物信息群里的老师同学,点评一下这篇文章 看看他的附图,结果好得不得了,以至于我不得不把所有附图也放上来 本文最大的问题就是生物信息分析不够专业,很多 流程都是常用软件加上默认参数,但不是最好的, 特别R语言画图,就是一坨屎,几种颜色分不开,我 600多度大眼睛,还要放大很多倍。 文章的分析稍微有些粗糙,他应该看一下,哪些基因表达变化的倍数与 染色体变化一致,也就是说是染色体变化导致的,还有另外一些基因 的表达变化远远高于低于染色体带来的变化,这要归结于其他原因 比如fusion gene,或者被调控等。 他这个技术最大的这个信息他居然没有充分利用。 文章叫做《GT-seq: parallel sequencing of single-cell genomes and transcriptomes》 macaulay2015.pdf nmeth.3370-S1.pdf 上面的想法,引起一个问题, 如果癌症都会有染色体级别的DNA变化, 必然导致基因表达改变,而且前者变化更大,很可能是主导因素,那么 我们现在大量的基因表达差异分析,又有什么意义呢? 反过来讲,这些染色体级别的DNA变化,又是谁引起的呢? 但是,即使把这些搞清楚了,又能怎么样?将来无非沿着两条腿走下去, 一个是继续寻找致癌机制,这个估计道路漫长啊,另外就是不管三七二十一, 通过检测找到某种癌症表现的共性,然后靶向某些基因,干死他, 后者思路更清晰,不过没那么简单。 由此想到,国内一大片跟着做下去的,无非继续检验了老外设备的技术可靠性。 其实,现在需要更多的临床信息,特别是药物与癌症互相作用等更多 其他方面的研究,对检测和治疗有直接帮助的东西,恰恰没有人做。 当前,医生们大多放弃了自己最有优势的病人资源,去和医学院生科院的 老师抢着杀小白鼠。 下半年嚷嚷的精准医疗,也不要盲目推出为好,无非就是一大堆外显子 测序,对于医院又能挣钱又发文章,对于病人呢,能改进治疗么,或者 根据突变的检测结果给予最恰当的药物匹配。 其实,几年前,我们就想做一个包括3000多种药物的库,与带有突变的 药物靶点进行匹配。几次基金不中,也就懒得多写这样的本子,还不够 浪费纸张的,有空可能会和公司合作,做一点有用的东西,希望能够 帮助一下癌症患者。
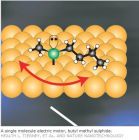
单分子阶梯事件分析 Single-molecule detection (SMD) and tracking in living cells is becoming a powerful method for the study of protein local environments, the time course of the enzymatic reaction, and the structure fluctuations of macromolecules, etc. Many single molecule studies have offered new insights into their localization, assembly and activation. Perhaps the dominant advantage of single-molecule fluorescence detection is that it can provide information on the spatial and temporal heterogeneity of molecule that underlies the ensemble average in conventional biochemical experiments. These spatial and temporal heterogeneities often appear as step events, for example, the discrete steps of photobleaching of single fluorescent molecules response to the number of subunits in membrane proteins. Hence , the step events analysis is becoming an important method of stochiometry study. However, SMD is also a difficult task. The most experiments of single-molecule fluorescence detection are approaching the limits of optical detection, and that the raw experimental data are inundated with all kinds of noise, especially, the Poisson distributed photon shot noise. These steps are so dim that it is very difficult to distinguish them. To leach out the useful information from these noisy raw data is still a challenging task. Perhaps the simplest and most commonly used method for the analysis of step events of single molecule inundated in these noisy data is the thresholding method. When a single-molecule trajectory has sufficient contrast between states, thresholds can be applied to distinguish the states of the molecule. These thresholds are typically chosen manually and introduce subjectivity into the analysis inevitablly. Before thresholding, binning of the data is also required, and this limits the temporal resolution of the measurement to be 1 or 2 orders of magnitude lower than the photon count rate to overcome the effects of shot noise. To mitigate the effects of shot noise, some investigators have applied filters to the data prior to applying a threshold. This can substantially improve the time resolution of the experiment by mitigating some of the effects of shot noise, but there is still the difficulty associated with choosing a threshold. Recently, many methods, such as hidden Markov models, applications of information theory, photon statistics in the context of two colors, maximum likelihood and Bayesian inferential estimation of change points using Poisson statistics, wavelet correlation, and wavelet shrinkage have been developed and applied to single molecule data as a means of extracting more accurate information about the system under observation. Although direct, model-independent information theoretical approaches may also work well, especially when a kinetic model is inapplicable. HMM, which uses all the information from the data prior and posterior, has enjoyed wide applicability and success. However, this method needs a long sequence to train the HMM for the parameter extraction. Many important experimental data sets are far too short to satisfy this requirement, so that the algorithm will converge to a local maximum depending on the initialization of the emission probability distribution. Another problem of HMMs is the necessary prior knowledge of the number of the states.
膜蛋白分子扩散运动模型 从单分子追踪(SPT)技术获得的单分子轨迹中,可以计算单分子扩散的均方位移( MSD : mean square displacement ),一般说来,该均方位移是单分子三维扩散运动的函数,我们这里只介绍二维扩散运动(膜运动)的形式。 根据 MSD 的不同,一般使用五种运动模型来描述膜蛋白的运动形式,分别是: 1) 稳态模型,也就是在观察时间内,蛋白分子几乎没有运动,在数量上定义为扩散系数 D 小于 4.6X10 -12 cm 2 /s ,如图中的 A 所示。 2) 布朗运动,蛋白分子进行简单地布朗运动,这种情况下, MSD-t 曲线是一条直线,如图中的 B 所示。 3) 直线扩散运动,分子以一个固定的速度沿着某个固定的方向进行扩散运动,同时叠加一个随机的扩散运动,,如图中的 C 所示。 4) 受限运动,这时分子是在一个受限的区域内进行布朗运动。直观地说,当膜蛋白陷在某个膜区内时,就会发生这种扩散运动,比如由蛋白骨架设定的膜区,如图中的 D 所示。这种情况下,我们通过对 MSD 曲线的拟合,可以获得模糊系数 D 、运动速度和约束区间尺度等重要信息。 5) 障碍扩散,这时蛋白分子的扩散运动受到周围其它障碍物的影响,而随着障碍物的不同,这种扩散也呈现出不同的运动轨迹,如图中的 E 、 F 所示。 应该注意到,上述是基于统计意义上的五种蛋白分子扩散运动分类模型,而实际上,由于所处细胞环境的复杂性和不断变化,分子往往呈现几种运动模型的混合状态。
今天早上看到新闻里面介绍2010年的诺贝尔物理奖,颁给了对石墨烯做出突破性实验的 Andre Geim和 Konstantin Novoselov。 查了一下原始论文,那个石墨烯的制备实验竟然是那么的简单。 原创: Our graphene films were prepared by mechanical exfoliation (repeated peeling) of small mesas of highly oriented pyrolytic graphite 【science, 306, 666, Oct 22, 2004】 (这篇文章的页码也够运气的;据报道,那个方法在此之前也有人试过,不过由于没有使用合适的检测方式,不知道是否得到了单分子。) 细节在文章的补充材料:The starting material was 1-mm-thick platelets of highly -oriented pyrolytic graphite (HOPG). We used commercially available HOPG of grades ZYH ( www.ntmdt.ru ) and HOPG-1 ( www.2spi.com ) with m 100,000 cm 2 /V s at 4K. Using dry etching in oxygen plasma ( S1 ), we first prepared 5 m m-deep mesas on top of the platelets (mesas were squares of various sizes from 20 m m to 2 mm). The structured surface was then pressed against a 1- m m-thick layer of a fresh wet photoresist spun over a glass substrate. After baking, the mesas became attached to the photoresist layer, which allowed us to cleave them off the rest of the HOPG sample. Then, using scotch tape 【胶带纸】we started repeatedly peeling flakes of graphite off the mesas. 【 http://www.sciencemag.org/cgi/data/306/5696/666/DC1/1 】 ACS的新闻介绍: Yet in 2004, Geim and Novoselov, who was a postdoc at that time, worked out a surprisingly simple method for exfoliating little chips of graphite by folding adhesive tape against the crystals and peeling apart the tape repeatedly http://pubs.acs.org/cen/news/88/i41/8841news4.html 当然,真正重要的是石墨烯分子特有的物理性质和实际应用。不过,如果得到了那个纯的石墨烯单分子层,后面的实验很多实验室都能够做到。 那么,为什么那两个俄国佬能够把单个的石墨烯分子从石墨晶体中用 胶带纸 给剥出来呢? 这里面的化学知识好像中学老师都教过。石墨的晶体结构是由一层层的分子组成,每个碳原子与周围3个碳原子形成3个完全等价的共价键,键长为1.421A(0.1421纳米),这样碳原子还剩下一个pi电子,同一层所有等价的碳原子的pi电子就形成了一个向2维空间无限扩展的共轭体系。而上下两层分子之间没有化学键,靠的是一种非键相互作用【说了白说】,英文里面有个词叫做stacking, 即pi电子之间的弱的静电作用。到底如何弱,看看分子之间的距离就明白了。 下图显示石墨烯之间的距离大约是3.354A。考察分子间的势能函数,可以计算出这个距离的作用力非常小,也就是通常所说的范德华作用力( van der Waals force)。 【图片来源:(1)w ikipedia(2) http://acigjapan.com/graphite.html 】 从英国剑桥的小分子晶体库( CSD )里面可以找出一个结构(代码是 OVALEN,分子式为C34H14,末端的氢原子没有显示)。虽然和石墨的晶型不同,但是上下两个分子堆在一起和石墨也蛮象。从它的3维SPACEFILL模型可以看出那个距离差不多就是原子间的密堆积了。 http://rspa.royalsocietypublishing.org/content/220/1141/157.abstract 为了帮助理解,我从PNAS的一篇文章里面找出一个理论模拟的结果。下图显示的是两个甲烷分子的势能曲线受压力的影响。因为有氢原子连在碳原子上,那个最低能量点比石墨晶体中分子间距离略微大一点 。【在微观尺度,1nM=10A=10亿分之一米】 http://www.pnas.org/content/95/4/1552.full 那个距离又是啥概念?不知道有多少读者会感觉到那个尺寸其实在生物里面也是非常的普遍和重要。 在生物体系,分子之间的非键作用是至关重要的。这里面一个是极性原子之间的氢键,它控制着生物微观过程的方向【过于简化,不过基本上就是那么回事】,另外一个便是非极性原子之间的 疏水相互作用 (hydrophobic), 它使得生命现象成为可能【更玄乎了点,举2个例子,这种作用使得蛋白质折叠成稳定的3维结构,也成就了形形色色的生物膜】 下面图示的是一个非常重要的药物分子(抑制剂)在酶的活性中心的结构,图上标有3个药物分子和蛋白之间的碳碳非键相互作用距离,都在3.6A左右。 【 有读者知道那是什么药吗?】 DNA分子中上下碱基对之间的距离也是在那个尺寸。 这个疏水相互作用的表象便是非极性的原子(团)聚集在一起,最常见的非极性原子便是碳了。人们分析过蛋白质晶体结构中(饱和)碳碳原子之间的非键相互作用距离,统计数据表明,那个距离的分布在3.6A左右有个峰。这个距离和石墨晶体中分子间的距离是很接近的,不同之处大概来自原子周围环境的细微差别。 下图显示的是碳元素的3种理想模型,来自炸药奖的官方网站。里面有本次获奖作品的详细介绍。第一个是足球烯C60,是1996年的化学奖项目。第二个是纳米碳管,有不少人都认为它比石墨烯的工作更应该得奖。第三个就是石墨烯了。 http://static.nobelprize.org/nobel_prizes/physics/laureates/2010/sciback_phy_10.pdf
 标签: 单分子
标签: 单分子