

 精选
精选
71. 胚胎的不对称性是如何形成的?
How is asymmetry determined in the embryo?
题记:旋转的纤毛有助于胚胎分辨左右,但科学家们仍在继续寻找使一个相对均匀的球形细胞变成有头有尾、有前有后不对称胚胎的最初原因。
人类在自发的对称性破缺方面总能表现出浓厚的兴趣。那些不对称的事物究竟是如何从当初对称的状态中被产生出来,那里到底发生了什么不可逆的故事?比如今天看来不均匀的宇宙,据说来源于一个对称奇点的爆炸;比如高温下无序而没有磁场的物质,温度降低后自发产生了有方向的磁场;比如本来没有形状均匀的水,温度下降后变成了固态规则的冰;再比如所有的人都开始于一个球对称的细胞,这些都是所谓的对称自发破缺的结果,也就是本来一个对称的体系(数学上对称)却发生了物理上的不对称的结果,而这个问题在发育生物学领域就是:在个体发育过程中,是否有共同的规则来决定不对称的方向(对称性破缺),从而产生不对称的个体? 为了明确这种空间的不对称,我们先来预习一下图1所示的解剖学上的专有名词。
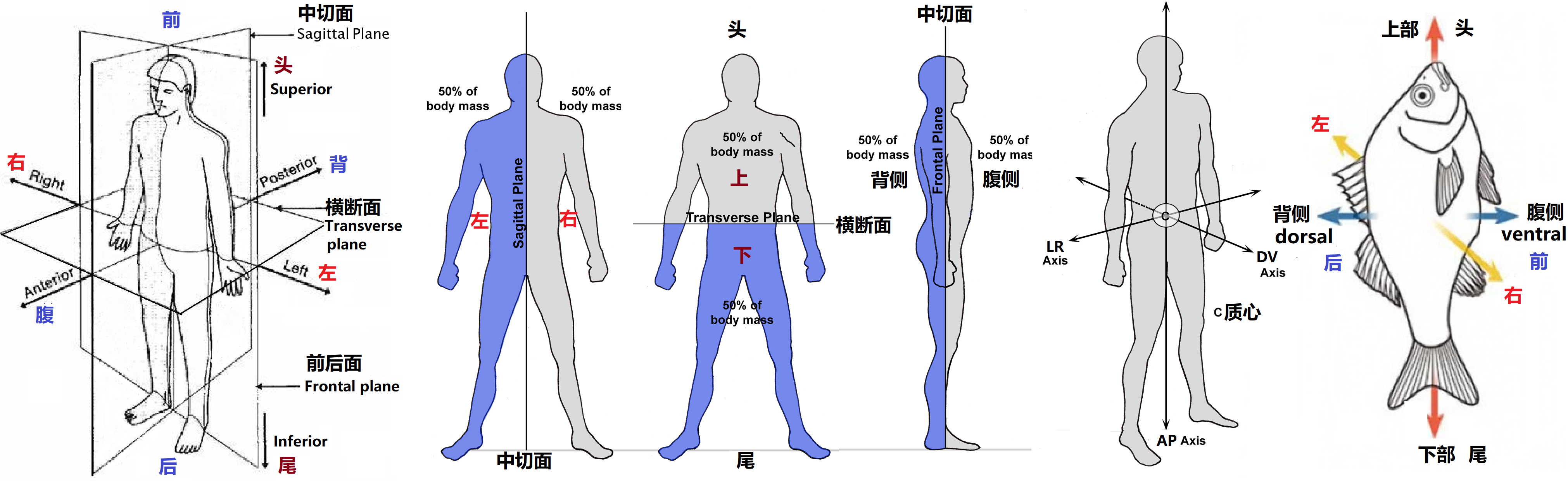
图1 生物体空间划分和名词规范
生物体有三个相互垂直的几何对称轴,如图1所示,这三个主轴是由三个互相垂直面的交线构成的,分别称为前后轴(AP)、背腹轴(DV)、左右轴(LR)。对人体而言,个体外部体征具有明显的左右对称性,即具有相对于中切面(Sagittal Plane)的镜像对称性,但人体的腹背和前后(头尾)显然都是不对称的。而且即便对于左右对称,人体内部器官的分配和器官组织(如心脏、肝脏、脾脏和肠道)的不对称也会打破人体左右的外部对称性。所以在胚胎发育过程中,通过复杂的遗传和表观遗传的级联反应,内脏器官的左右不对称放置是脊椎动物身体形态的主要特征。虽然大多数生物体都具有和人相似的左右对称结构(如图1中的鱼),但它们的胚胎发育却都是从一个对称度高的细胞开始逐渐分裂成长而来,最后都会发生对称性破缺变成如图2所示的不对称的胚胎囊胚(blastocyst)。大量的研究表明不同脊椎动物左右对称形成的机制既有共性,也有差异,而且左右不对称的建立与腹背轴(anterior–posterior axis)对称伸长之间还存在某种关联。无论如何,发育生物学家一直希望找到这种不对称起源的本质,而这个问题的答案会让人类理解和掌握发育的基本生物学过程,从而为解决发育缺陷和遗传形态综合症(歧形或异位症)提供有效的治疗方案。
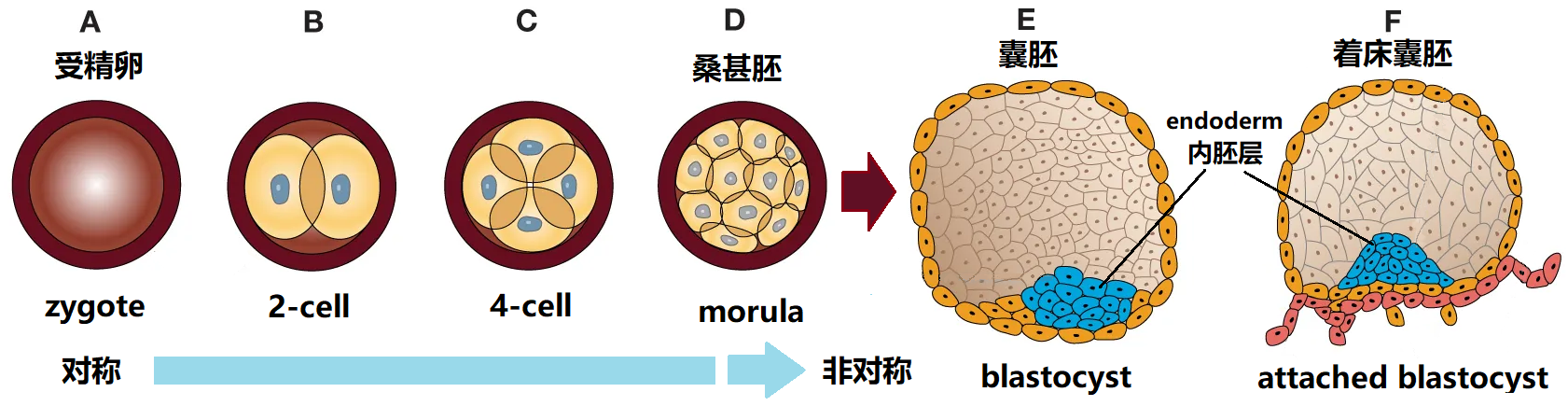
图2 胚胎前期囊胚的形成过程
对于生物体不对称的出现,第一个问题自然是:个体的不对称方向是由基因、环境造成,还是由偶然因素决定的? 答案当然会首先否定偶然因素,因为偶然因素只会导致随机的结果,但生物体存在对称性而且不对称也有惊人的一致性(如人都是心脏偏左的)。所谓的偶然因素实际上是指发育过程中“随机噪声”所导致的波动不对称性,但它显然不是造成生物体不对称发育的主要原因,如果是,那所有的对称性都应该不会出现,但胚胎的左右对称却依然在“噪声”中被保持了下来。第二种认为不对称出现的原因是后天的使用或特别加强的神经活动,然而这也肯定不是主要原因。招潮蟹有一个钳子很大,但当它失去大钳子的时候,新长出的钳子依然会大于那个用了更久的小钳子。显然生物体不对称的方向是由基因和环境两种因素共同决定的,基因产生了某种蛋白质的表达结果,而环境影响了这种表达的过程和发展方向。

图3 胚胎形成过程中的不对称分裂
显然生物个体的不对称性在其胚胎形成之前就已经建立(如图2所示),胚胎后期的发育过程显然只是由小到大的过程。然而胚胎的形成都是从对称分裂的细胞群开始,后来才逐渐发育成前后左右不对称的生物个体,也就是脊椎动物的囊胚从一个受精卵开始逐步形成具有前后轴(AP)、背腹轴(DV)、左右轴(LR)都不对称的生命体征。在囊胚的发育过程中,一系列不对称的细胞分裂对于建立胚胎三个主轴(AP、DV、LR)及其对称破缺至关重要,见图3所示。首先来源于精子的中心体会启动皮层肌动球蛋白网络(cortical actomyosin network)的运动,从而导致PAR(protease activated receptors,蛋白酶激活受体)蛋白的极化分布。然后PAR蛋白介导了前卵裂球AB和后卵裂球P1的不均等分裂,在四细胞阶段开始建立DV轴(腹背轴),P1的两个子代P2和EMS以及AB的子代ABa和ABp之间发生特定的细胞间相互作用,在此后不久LR轴由ABa和ABp的之后的分裂模式建立。
尽管卵细胞的不对称分裂导致了沿不同对称轴的破缺,但左右对称的选择又是什么导致的?显然囊胚的AP轴(头尾的方向)和DV轴(腹和背的方向)可以通过外部因素(重力和精子的进入点)来设定,但囊胚似乎没有独立的方法来选择左右方向,因为没有已知的宏观因素可以区分左和右而进行左右不对称的细胞分裂。也就是在没有任何化学或物理的宏观特征来区分左和右的情况下,LR轴如何能够始终如一地朝向前后(AP)轴和背腹(DV)轴? 1963年赫胥黎和德比尔提出了一种可能的方法,即可以使用基本力来确定LR轴相对于其他两个轴的方向。他们提出,在胚胎发育过程中,脊索不对称是通过电流沿着脊索的长度方向(前后)进行定向的,如果在脊索的背侧或腹侧测量,电流会产生指向L或R的磁场力。虽然已经观察到地球地磁场反转和贝壳手性之间的相关性(Harrison和Funnel, 1964),但这种因果关系的本质依然是未知的,并且迄今为止没有确切证据表明地球磁场在物种LR选择中被利用。
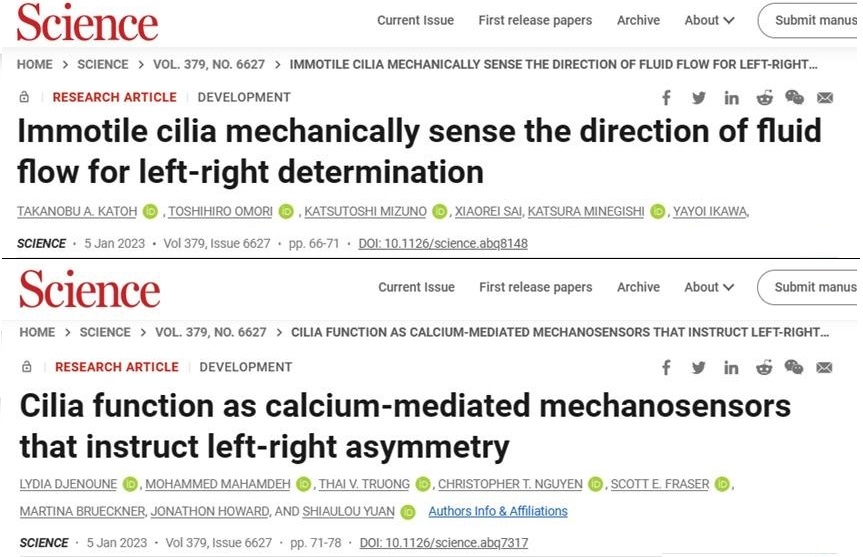
图4 科学杂志的两篇论文截图
不是磁场力那会是什么力?机械螺旋力。2023年1月《科学》杂志背靠背发表了两篇论文来解释这个问题,如图4所示。这两篇论文都给出了确定左右对称方向和产生左右非对称的源头:一种具有螺旋方向的绒毛。在大多数脊椎动物中,左右的差异在早期胚胎发生时由一小群称为左右组织(left-right organizer)的细胞所指定。在这个组织体内,可运动的纤毛迅速移动,产生细胞外液体向左方向流动,这是左右差异的第一个迹象,但这种流动是如何被感知和转导到后来的分子和解剖上的左右不对称的,目前还不清楚。通过对小鼠胚胎的研究,Katoh等人发现,静止的纤毛能够感知由液体流动所产生的机械力,并提出了一种感知液体流动方向的生物物理机制。Djenoune等人利用光学镊子和实时成像技术,在斑马鱼身上进行了独立研究,发现组织体中不动的纤毛具有机械传感器的功能,可将细胞外液体流动转化为钙信号。当活动纤毛瘫痪,正常体液流动停止时,对纤毛的机械操控可以挽救甚至逆转胚胎左右生长的模式。因此,纤毛对流体力的感知是胚胎左右偏好对称破缺所必要的结果。
当然以上的两篇论文给出了左右区分的发展机制,但远远不是这个问题的最终答案,对于生物体左右偏好这个问题最为根本的原因现在依然存在多种解释,主要有两种基本的对称破缺的解释模型,一种认为导致对称破缺的对称偏好来源于生物分子本身就存在的手性对称性,而另外一种则认为是发育过程中信号的非对称表达,如下图5所示。
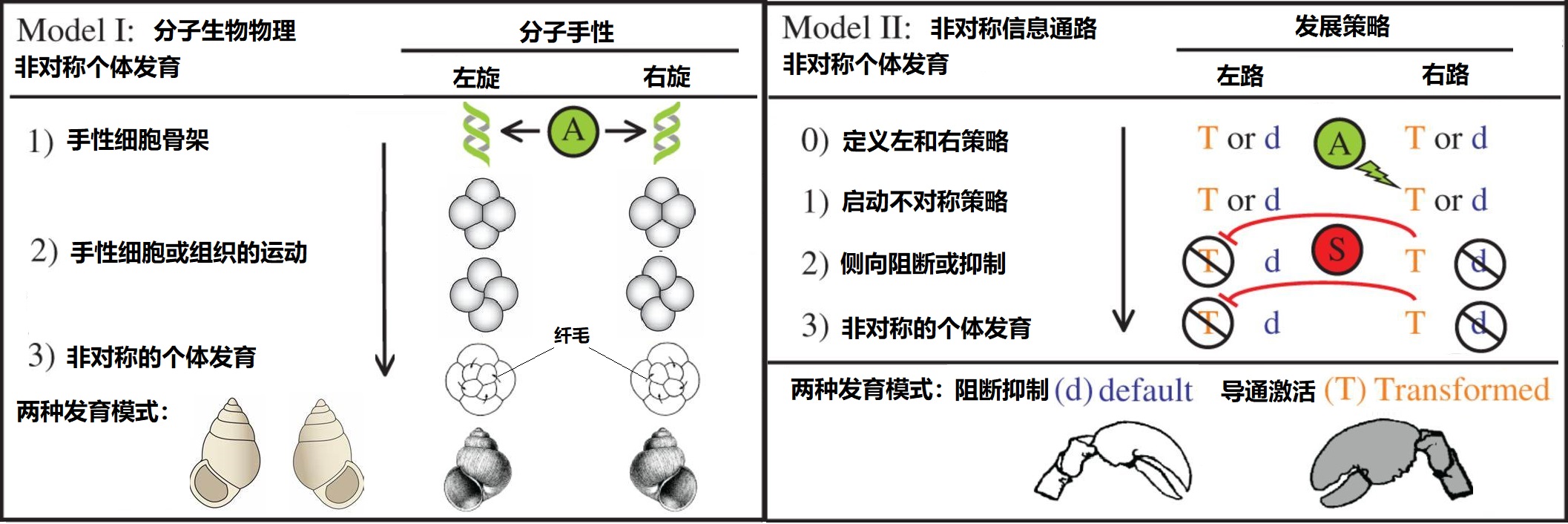
图5 解释对称偏好的主要模型[文献1]
然而无论如何问题仍然多于答案,最后我们依然还有很多问题要问:为什么生物体需要或存在不对称?不对称的心脏、肠道和大脑对结构和生理有什么影响?为什么所有正常的个体不仅不对称,而且不对称的方向也是相同的(比如人为啥都是心脏偏左的不对称,而不是心脏偏左和偏右的各占一半,内脏都反方向的镜像人应该不存在生理差异)?在进化过程中,手性不对称是什么时候出现的?在定向不对称出现之前,是否存在真正的双向对称生物(Cooke, 2004)?它是否与较低形式的手性有关(如蜗牛壳盘绕和某些植物的手性)?脊椎动物胚胎的不对称是在哪个发育阶段开始的?要回答这些问题,需要在分子、遗传和生化层面上对胚胎中不对称偏好的形成都有详细的了解。
转载本文请联系原作者获取授权,同时请注明本文来自张林科学网博客。
链接地址:https://m.sciencenet.cn/blog-318012-1419409.html?mobile=1
收藏