博文
时间信息的贝叶斯优化知觉
||
时间信息的贝叶斯优化知觉
陈有国**1,2 彭春花3 刘培朵1,2 余婕1,2
(1西南大学心理学部时间心理学研究中心,重庆,400715)
(2西南大学心理学与社会发展研究中心,重庆,400715)
(3重庆文理学院重庆市情绪与心理健康重点实验室,重庆,402160)
本文发表于《心理科学》2024年第1期见:http://www.psysci.org/CN/10.16719/j.cnki.1671-6981.20240102
摘要:大脑运用贝叶斯推理加工时间信息形成优化知觉。贝叶斯模型在趋中效应、多源时间信息整合、时空干扰效应以及同时性的贝叶斯校准这四个课题上取得了较大的进展。本文在测量-估计-决策三阶段模型的框架下,对以上课题中的贝叶斯模型进行分析。未来需检验不同形式的时间信息先验的合理性,厘清似然性的心理表征,探明时间信息贝叶斯估计的神经基础。应该结合时间信息加工模型与贝叶斯模型的优势,为时间信息加工的认知神经机制研究提供新的思路。
关键词:时间信息、贝叶斯推理、趋中效应、时间信息整合、时空干扰效应、同时性的贝叶斯校准
贝叶斯理论在认知科学中得到了广泛的应用。大脑能够表征外部世界的不确定性,并根据标准的贝叶斯推理,将信念(或经验)与新输入的信息进行整合,做出最优决策(Pouget et al., 2013)。大脑在不同类型的心理加工中使用贝叶斯推理,包括感知、记忆、语言、运动控制、社会认知等(Rescorla, 2021)。贝叶斯理论也在“毫秒至秒”范围的时间知觉研究(Miyazaki et al., 2006; Shi et al., 2013)以及“分至年”范围的日常生活事件的时间估计研究(Griffiths & Tenenbaum, 2011)中得到了广泛的应用。
时间知觉(time perception)是人类生存必须的基本能力,指毫秒至大约3秒内的时间信息加工。其研究对象为基本的时间经验,包括同时性(simultaneity)、非同时性(successiveness)、时间顺序(temporal order)、主观时距(subjective duration)等(Pöppel, 1997)。同时性知觉是个体判断两个感觉事件同时发生的知觉。非同时性(或相继性)知觉指个体判断两个事件不是同时发生,但不能判断它们的先后顺序的知觉。时序知觉是对顺序性的知觉,关注人类如何判断两个事件出现的先后顺序。时距知觉是对持续性的知觉,关注人类如何在时间维度上累加时间单元(如脉冲、神经振荡),形成对事件持续时间的知觉。经典的时间信息加工模型和贝叶斯推理存在相似之处(图1)。时间信息加工模型包括时钟、记忆和决策三个阶段,时钟阶段由节拍器、开关和累加器组成;记忆阶段由参照记忆和工作记忆组成;决策阶段比较参照记忆和工作记忆中的时间信息,并做出反应(Gibbon, 1977)。Shi等(2013)指出时钟、参照记忆、工作记忆以及决策阶段,可以分别类比于贝叶斯推理的似然性(likelihood)、先验(prior)、后验(posterior)和损失函数(loss function)。在时钟阶段,假设外部输入的时距信息为D,累加器测量节拍器发出的脉冲数量为S。由于神经系统存在噪音,脉冲数量的似然性为概率分布p(S|D)。在记忆阶段,以前对时距进行估计的先验经验p(D)储存于参照记忆中。根据贝叶斯定理,大脑对先验p(D)和似然性p(S|D)进行整合,计算主观时间的后验:p(D│S)=(p(S|D)p(D))/(p(S))。在决策阶段,损失函数L是每种行为对应的损失,个体根据后验和损失函数对时间信息进行优化估计(Shi et al., 2013)。贝叶斯观点认为时间知觉是无意识推理的结果,称之为时间信息的贝叶斯优化知觉(Jazayeri & Shadlen, 2010; Shi et al., 2013)。
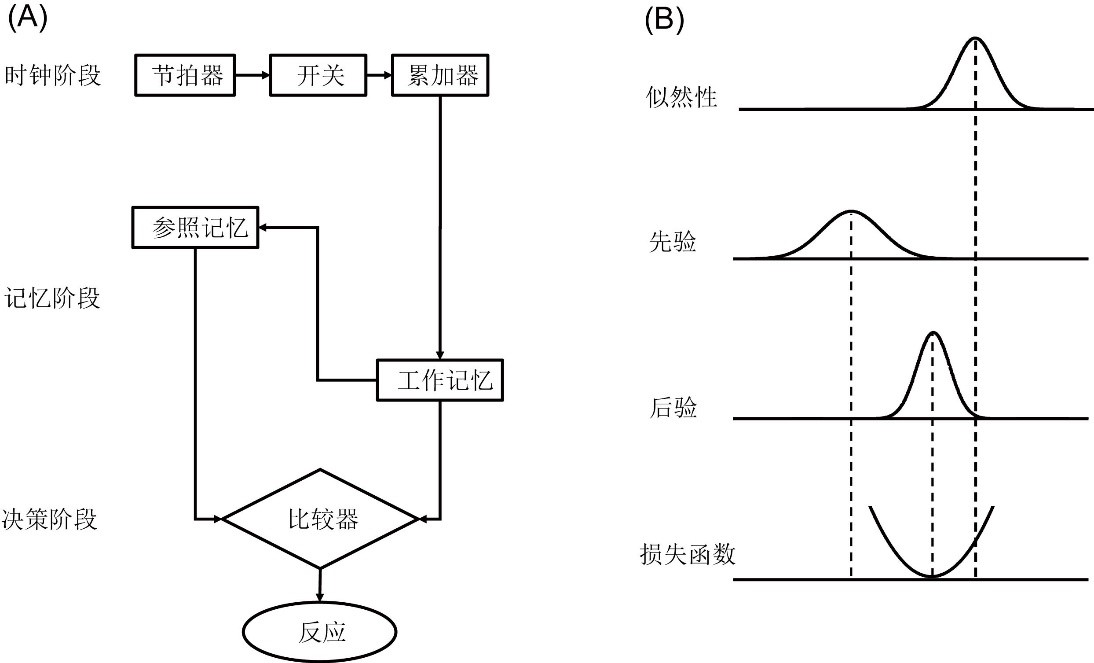
图1 时间信息加工模型与贝叶斯推理。(A)经典的时间信息加工模型。(B)典型的贝叶斯推理过程。修改自Shi et al., 2013。
贝叶斯理论在趋中效应、多源时间信息整合、时空干扰效应以及同时性的贝叶斯校准(Bayesian calibration of simultaneity)这四个课题上取得了卓有成效的进展。趋中效应与同时性的贝叶斯校准涉及时间背景对时距、时序信息加工的调节;多源时间信息整合涉及相同感觉通道内或者不同感觉通道的时间信息整合;时空干扰效应涉及时间信息与空间信息的整合加工。对以上四个课题的贝叶斯模型进行分析,可以了解贝叶斯模型在时间信息加工领域应用的基本思路与最新进展。本文不仅对时间信息加工领域的研究有重要的启示,而且有助于其他认知科学领域的研究者了解运用贝叶斯理论对心理过程进行建模的基本方法。
1 趋中效应
趋中效应(central tendency effect),也称为Vierordt定律。被试按键复制一系列时距,会出现高估短时距低估长时距,即长时距和短时距的估计值都向中间趋近。Jazayeri 和 Shadlen(2010)构建了贝叶斯最小平方模型(Bayesian least square model,BLS)解释时间知觉的趋中效应,该模型包括三个阶段(图2A)。(1)测量。给被试呈现一系列时距中的某一物理时距ts,大脑测量该时距的似然性为p(tm|ts)。(2)估计。在Jazayeri 和 Shadlen(2010)的研究中,每个时距的概率相同,服从均匀分布。假设被试会学习到该均匀分布,将之作为先验p(ts)。大脑通过贝叶斯定理整合先验和似然性,计算出后验: p(t_s│t_m )=(p(t_m│t_s )p(t_s))/(p(t_m)) 。令te为后验的最优估计值,采用平方误差损失函数L(t_e,t_s)=〖(t_e-t_s)〗^2进行最优估计。根据后验和损失函数可以计算期望损失:E(L)=∫▒〖L(t_e,t_s)〗 p(t_s│t_m )dt_s。将期望损失最小化,得到的最优估计值te为后验均值。由似然性均值至后验均值,会产生趋近先验中心的偏差,出现时距知觉的趋中效应(图2B)。(3)产生。被试根据最优估计值te按键产生时距tp为正态分布p(tp|te)。BLS认为被试能够学习到外部刺激的统计结构作为先验,但并未考虑先验从无到有的学习更新的过程。

图2 贝叶斯最小平方模型。(A)正态分布似然性的三阶段贝叶斯模型。(B)贝叶斯推理过程。
迭代贝叶斯估计模型(iterative Bayesian estimation model,IBE)认为先验会随着个体的学习过程不断迭代更新 (Petzschner & Glasauer, 2011; Petzschner et al., 2015)。运用三阶段贝叶斯模型对IBE进行介绍(图3A)。(1)测量。根据韦伯-费希纳定理,对于某一物理时距ts,大脑测量该时距的似然性服从对数正态分布(log-normal distribution)p(tm|ts)。将该似然性进行对数转化,得到对数尺度的似然性为正态分布p(Sm|Ss),其均值为S_s=ln(t_s)。(2)估计。被试根据以前的估计形成的先验为正态分布p(Sτ)。根据贝叶斯定理计算出后验:p(S_τ│S_m )=(p(S_m |S_τ)p(S_τ ))/(p(S_m)),其最优估计Se为后验均值。个体在每次贝叶斯估计时,其先验也会遵循卡尔曼滤波原则进行更新(图3B)。在第i次试验,先验均值S_(prior,i)为第i-1次似然性均值与i-1次先验均值的加权平均值S_(prior,i)=k_(i-1)∙S_(s,i-1)+(1-k_(i-1))∙S_(prior,i-1);后验均值为似然性均值与先验均值的加权平均值S_(e,i)=w_i∙S_(s,i)+(1-w_i)∙S_(prior,i)。(3)产生。运用指数函数将最优估计Se转化为估计时距t_e=e^(S_e ),并根据估计时距te按键产生时距tp为正态分布p(tp|te)。在第i次试验中,IBE模型的先验均值S_(prior,i)是由以前的似然性均值(S_(s,i-1),…,S_(s,1))的加权平均值构成的(图3B),所以S_(prior,i)会趋近于出现过的Ss的平均数。后验均值S_(e,i)是先验均值S_(prior,i)与新似然性均值S_(s,i)的加权平均值,所以S_(e,i)相对于S_(s,i)会向中心趋近。IBE模型采用了不断迭代更新的正态分布作为先验,避免了BLS模型必须人为设定先验分布函数(如均匀分布)的困境。
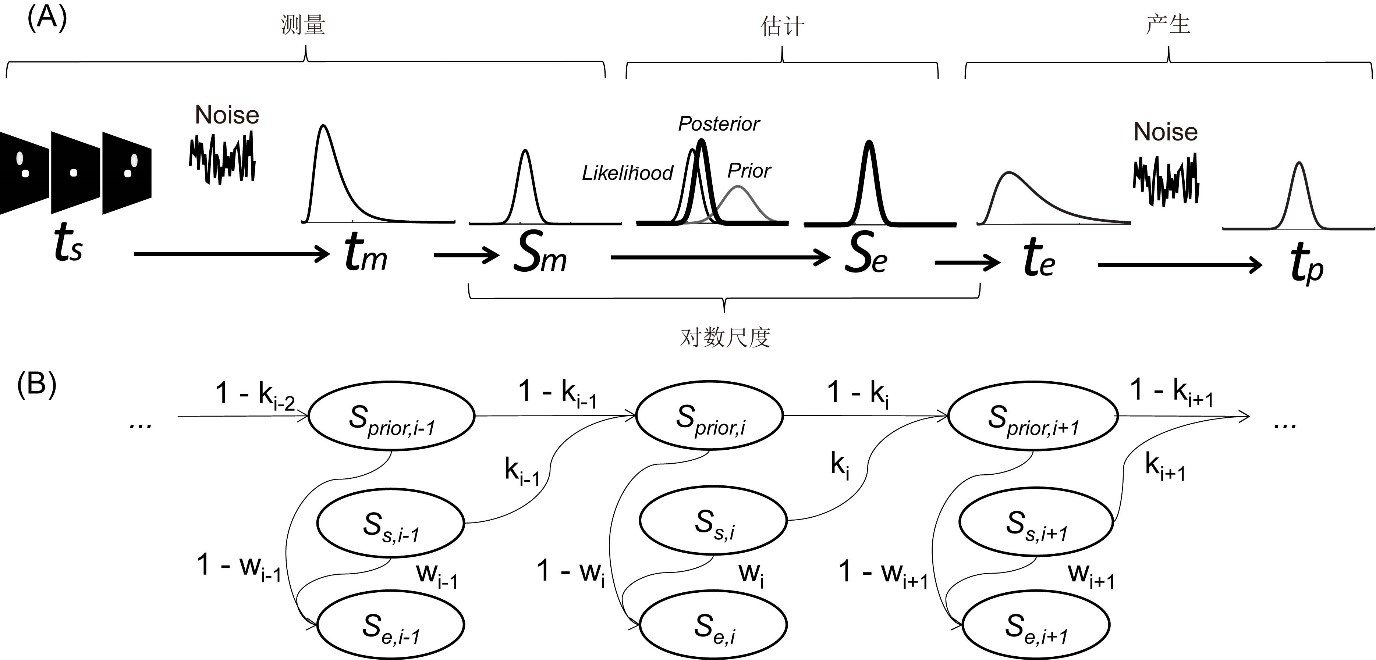
图3 迭代贝叶斯估计模型。(A)对数正态分布似然性的三阶段贝叶斯模型。(B)卡尔曼滤波迭代更新过程。
趋中效应的贝叶斯模型得到了不同类型人群数据的支持。贝叶斯模型预测时距表征的变异性越大,时距知觉越会受到先验的影响,则趋中效应越强;反之,时距表征越精确,则趋中效应越弱(Jazayeri & Shadlen, 2010)。与贝叶斯模型预测一致,儿童、老年人和帕金森病患者的时距表征的变异性较大,故这三类人群的趋中效应较强(Gu et al., 2016; Hallez et al., 2019);鼓手的时距知觉较为精确,故其趋中效应较弱(Cicchini et al., 2012)。有研究报告自闭症儿童的趋中效应弱于典型发育儿童(Karaminis et al., 2016)。根据贝叶斯理论,可能存在两种机制调节自闭症患者的趋中效应强度。第一种观点认为自闭症患者会自上而下(top-down)地抑制先验信息,在贝叶斯估计中更加依赖输入信息的似然性,导致其趋中效应更弱(Karaminis et al., 2016; Pellicano & Burr, 2012)。第二种观点认为自闭症患者的知觉加工同时存在自上而下与自下而上(bottom-up)的调节机制,其中自下而上的机制让自闭症患者减少感觉加工噪音(即降低似然性的变异性,增强时间感受性),同样会让自闭症患者的趋中效应更弱(Brock, 2012)。Casassus等(2019)对时间知觉与自闭症的关系进行了系统综述,发现无论是儿童还是成年人,不同研究报告的自闭症患者与正常人的时间感受性的关系并不一致 (Casassus et al., 2019)。未来可从年龄发育的角度,考察上述两种机制调节自闭症患者趋中效应强度的不同趋势。
2 多源时间信息整合
相同通道内以及不同感觉通道的时间信息均可以发生相互影响 (Burr et al., 2013; Chen et al., 2018)。最大似然估计模型(maximum likelihood estimation, MLE)和因果推论模型(causal inference model)对多源时间信息整合进行了解释。
MLE模型认为大脑运用贝叶斯定理整合不同来源的感觉信息,增加估计的可靠性(图4A)(Ernst & Banks, 2002; Pouget et al., 2013)。该模型假设先验为均匀分布p(S),大脑测量输入的各个感觉信息后得到的似然性为p(S_1,⋯,S_i |S)。根据贝叶斯定理计算后验:p(S│S_1,⋯〖,S〗_i )=(p(S_1,⋯〖,S〗_i |S)p(S))/(p(S_1,⋯S_i))。其估计值S ̂为各个感觉信息的加权平均值:S ̂=∑▒〖w_i S ̂_i 〗,其中w_i=(1⁄(σ_i^2 ))/(∑▒1⁄(σ_j^2 )),σi为第i个感觉信息的标准差。有研究将MLE应用于听触时距知觉,并证实听触时距的最优估计值位于听觉时距和触觉时距长度之间,并且最优估计标准差σ_AH=(σ_A σ_H)/√(σ_A^2+σ_H^2 )要小于单独的听觉时距和触觉时距的标准差(σA、σH)(Wing et al., 2010)。
因果推论模型最初被用于判断多重感觉信息是否来自于相同的位置(Körding et al., 2007),后来被用于判断不同的感觉信息是否来自于相同的时间点(Elliott et al., 2014)。假设外部输入的时间信息为tA和tB, 被试需推断它们是属于相同源S(记为C=1), 还是不同的源SA和SB(记为C=2)。该模型可以用层级贝叶斯模型进行描述(图4B),并分解为相同源(C=1)和不同源(C=2)两个简单的贝叶斯模型(Shams & Beierholm, 2010)。当时间信息来自于同一源时(C=1,图4C),因果推论模型的先验为正态分布p(S)。外部输入的两个时间信息的似然性分别为p(t_A |S)和p(t_B |S)。将先验和两个时间信息的似然性整合在一起形成唯一的后验p(S|t_A,t_B)。当时间信息来自于不同的源时(C=2,图4D), 先验分布p(S_A )p(S_B)与两个时间信息的似然性p(t_A |S)和p(t_B |S)分别整合在一起, 形成两个后验p(S_A,S_B |t_A,t_B)。
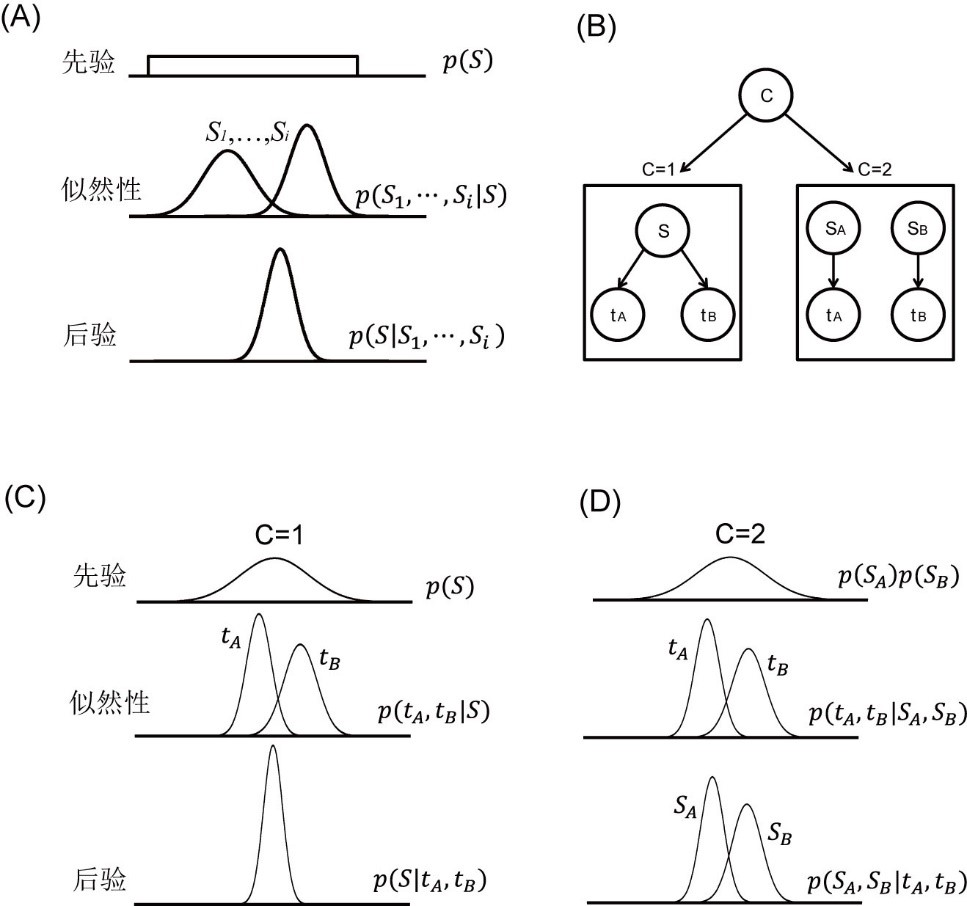
图4最大似然估计模型与因果推论模型。(A)最大似然估计模型。(B)层级因果推论贝叶斯模型。(C)相同源与(D)不同源条件下的因果推论模型。
多源时间信息整合研究揭示了不同类型人群感觉信息加工的特点。大约8岁的儿童不能有效地整合视听时间信息,而采取了亚优化(sub-optimal)的转换策略,在不同的试验中会单独根据视觉或者听觉时距进行反应;9至12岁的儿童只进行了部分整合(Adams, 2016)。恢复听力的耳聋儿童会整合视觉与听觉信息促进有效判断,其视听时距的辨别能力显著强于单独的视觉与听觉时距辨别能力(Gori et al., 2017)。精神分裂症患者和自闭症患者的视听时间信息整合能力受损,该整合能力与幻觉和社交障碍关系密切(Zhou et al., 2018)。
3 时空干扰效应
时间和空间信息加工会相互干扰。Kappa效应和Tau效应是最典型的两种时空干扰效应。Kappa效应涉及空间信息对时间知觉的影响,依次呈现两次闪光,如果固定两次闪光的时间间隔,会发现闪光之间的空间距离越长,知觉到的时间间隔越长(Cohen et al., 1953)。Tau效应涉及时间信息对空间知觉的影响,如果固定两次闪光之间的空间距离,会知觉到闪光之间的空间距离随着时间间隔的增加而增加(Helson, 1930)。
早期研究认为Kappa效应源自内隐运动,即个体会将不同位置相继出现的闪光,理解为以恒定的速度进行运动的同一物体。Jones 和 Huang(1982)基于恒速假设构建了定量解释Kappa效应的恒速模型。该模型提出个体知觉到的刺激间时间间隔te是实际时间ts和期望时间的加权平均值,其中期望时间为刺激间的空间距离l与恒定速度v0的比值,即t_e=ωt_s+(1-ω) l/v_0 。
近期研究表明Kappa效应是大脑运用贝叶斯定理进行无意识推理的结果。Chen等(2016)采用三阶段贝叶斯模型对恒速模型进行了重新解释(图2A)。(1)测量。给被试呈现某一物理时距ts, 该时距的似然性是正态分布p(tm|ts),其均值为ts,标准差为σm。(2)估计。令两次刺激之间的距离为l,恒定速度为v0,则先验为正态分布p(τ),其均值为l/v_0 ,标准差为στ。根据贝叶斯定理计算出后验:p(τ|t_m)=(p(t_m |τ)p(τ))/(p(t_m))。后验估计值为t ̂_e=ωt_s+(1-ω) l/v_0 ,其中ω=(σ_τ^2)/(σ_τ^2+σ_m^2 )。该表达式与Jones 和 Huang(1982)提出的恒速模型的表达式完全一致。(3)产生。根据估计时距te按键产生时距tp为正态分布p(tp|te)。Chen等(2016)发现该模型不能解释Kappa效应的减速趋势,即随着刺激间距离的增加,知觉到的时距并非线性增加,其增加的速度会逐渐减缓。
Chen等(2021)将韦伯-费希纳定律与恒速模型整合,提出对数版恒速模型。该模型加工阶段与IBE模型的贝叶斯估计过程一致(图3A)。(1)测量。大脑首先对物理时距ts进行测量并得到tm,其概率分布为对数正态分布p(tm|ts),然后采用对数函数将tm转化为对数尺度的内部时间Sm,其似然性为正态分布p(Sm|Ss)。(2)估计。在对数尺度下,先验为正态分布p(Sτ),其均值为l/v_0 的对数转化值。根据贝叶斯定理计算后验:p(S_τ |S_m)=(p(S_m |S_τ)p(S_τ))/(p(S_m))。最优估计值为后验均值Se。(3)产生。运用指数函数将最优估计值Se转化为估计时间te。根据估计时间按键产生时距tp为正态分布p(tp|te)。对数版恒速模型认为减速趋势是由于韦伯-费希纳定律导致的,其对Kappa效应行为数据的拟合度优于原版恒速模型(Chen et al., 2021)。
4 同时性的贝叶斯校准
时序判断的主观同时点会受到先前时序判断的调节,符合贝叶斯模型的预测,称之为同时性的贝叶斯校准。Miyazaki等(2006)将电极安置于被试左右手的食指上,并分别给予3毫秒电脉冲刺激。令右手电脉冲先于左手出现记为正时间条件,反之为负时间条件。该实验包括两种时间背景条件:正80毫秒条件的时序信息从均值和标准差均为80毫秒的正态分布中抽样;负80毫秒条件的时序信息从均值为-80毫秒,标准差为80毫秒的正态分布中抽样。同一组被试需要完成两种时间背景下的时序判断任务,结果表明时间背景调节时序判断的主观同时点,正80毫秒条件的主观同时点为-41毫秒,负80毫秒条件的主观同时点为56毫秒。
Miyazaki等(2006)构建了贝叶斯模型来解释时间背景如何调节时序知觉。三阶段贝叶斯模型如下:(1)测量。给与被试呈现一对电脉冲作为时序信息,该时序信息的似然性为正态分布p(tsensed|ttrue)。(2)估计。假设被试将所有时序信息的统计分布作为先验p(ttrue)。根据贝叶斯定理计算出后验:p(t_true |t_sensed)=(p(t_sensed |t_true)p(t_true))/(p(t_sensed))。(3)决策。根据信号检测论,后验分布中估计时间大于0的概率对应于被试判断“右手先出现电脉冲”的概率P(testimated>0)。以正80毫秒条件为例,先验为两次电脉冲时间间隔的统计分布,其均值和标准差均为80毫秒(图5)。图中的a、b、c、d分别表示呈现给被试的4个时序信息,其中a和b为负时序信息(左手先出现电脉冲),c和d为正时序信息。由于c的均值与先验的均值相同,故c的后验均值不变;a、b和d的后验均值均向先验均值(80毫秒)趋近。在判断概率阶段,被试选择“右手先出现”的比率为后验分布中时间大于0对应的概率(图5中阴影的面积)。采用累加正态分布函数对被试选择右手先出现的比率进行拟合(图5中的S型虚线),可知正80毫秒条件的主观同时点为负值。
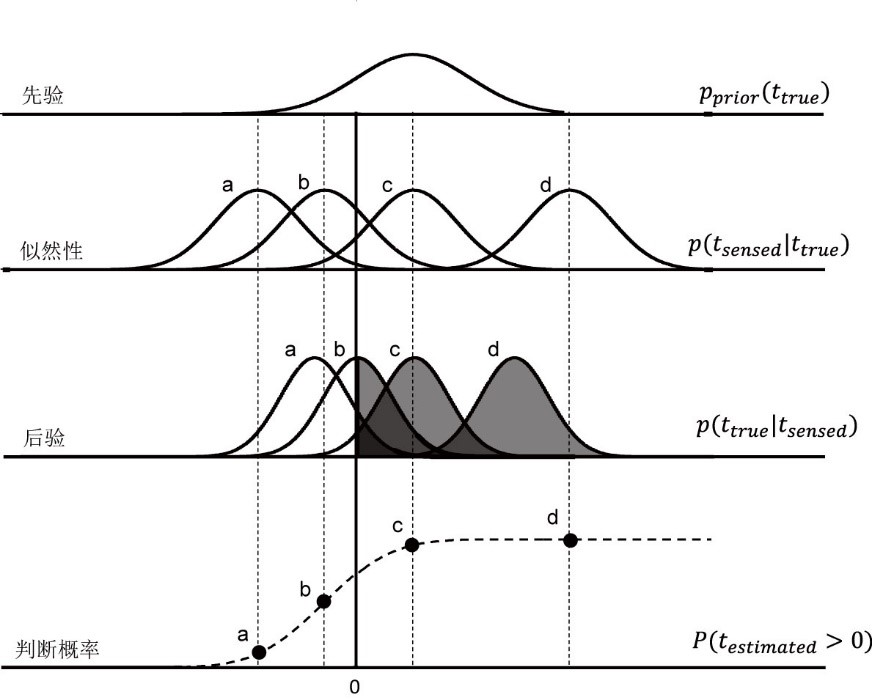
图5 时间背景调节时序知觉的贝叶斯模型
贝叶斯模型已被应用于研究自闭症患者的时序知觉。Wada等(2023)发现自闭症患者完成触觉时序判断任务的标准差与正常人群并没有显著差异,但自闭症患者进行触觉时序判断的主观相等点的偏差更小。该结果提示自闭症患者抑制了先验信息,支持自上而下的调节机制(Pellicano & Burr, 2012)。也有研究报告自闭症患者的触觉时序判断的标准差小于正常人群,即自闭症患者降低了感觉加工噪音,支持自下而上的调节机制(Brock, 2012; Ide et al., 2019)。未来尚需进一步的研究理清两种调节机制与自闭症患者时序知觉的关系。
5 总结与展望
贝叶斯理论为时间知觉研究提供了全新的视角。趋中效应、多源时间信息整合、时空干扰效应以及同时性的贝叶斯校准均可以采用测量-估计-决策三阶段贝叶斯模型加以描述(图6)。(1)测量。研究者根据韦伯定律或者费希纳定律构建输入时距的似然性。根据时间信息加工模型,时距的内部表征为正态分布,其标准差会随着其平均值的增加而增加,称之为标量变异(scalar variability)(Gibbon, 1977; Jazayeri & Shadlen, 2010)。标量变异是韦伯定律(感觉刺激的最小可觉差与刺激的强度的比值为一常数,∆I⁄I=k)在时间信息加工领域的具体表现。有研究者根据韦伯定律假设时距的似然性为正态分布,并根据标量变异对这些时距的标准差进行建模σ_m=w_m t_s(图2)(Chen et al., 2016; Jazayeri & Shadlen, 2010)。费希纳根据韦伯定律推导出费希纳定律(心理感觉正比于刺激强度的对数,S∝lnI)。韦伯定律和费希纳定律统称为韦伯-费希纳定律(Billock & Tsou, 2011)。有研究者根据费希纳定律假设时距的似然性为对数正态分布,该似然性(tm)经过对数转化,成为对数尺度下的正态分布似然性(Sm,图3A)(Chen et al., 2021)。(2)估计。个体根据日常生活的经验或者在实验室经过学习形成先验,按照贝叶斯定理整合先验和似然性得到后验。(3)决策。根据后验与损失函数,计算最优估计值。最小平方和最大后验是贝叶斯模型常用的两种最优估计策略。最小平方采用平方误差损失函数,其最优估计为后验的平均数;最大后验则认为最优估计对应于后验的众数(Jazayeri & Shadlen, 2010)。个体在决策之后产生行为反应。时间复制任务采用正态分布对被试的产生时距进行建模(Jazayeri & Shadlen, 2010);时间比较任务则采用信号检测论对被试选择“比较时距更长”的比率进行建模(Chen et al., 2021)。未来对时间信息的贝叶斯优化知觉的研究需要回答以下几个问题。
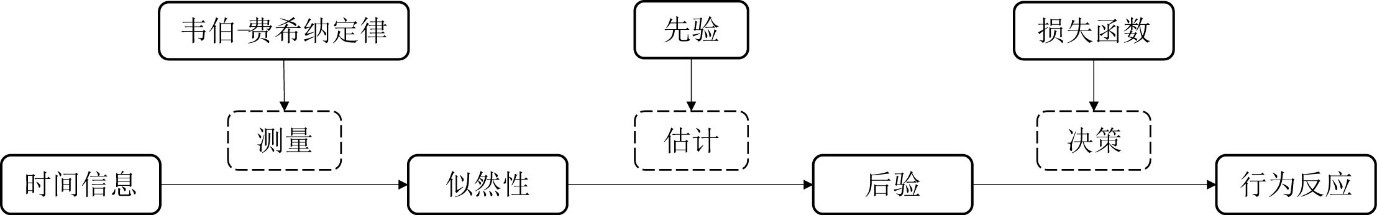
图6 三阶段贝叶斯模型图示
第一,检验不同形式的时间信息先验的合理性。在趋中效应研究中,各个时距的出现概率相同,时间信息的统计结构较为简单(离散的均匀分布),但研究者对于该效应中先验的心理表征还未达成一致。Roach等(2017)假设个体经过少量的学习便可以获得近似的正态分布作为先验;Jazayeri 和 Shadlen(2010)认为经过充分的学习,个体能够获得均匀分布作为先验;Acerbi等(2012)采用非参数方法根据行为数据重构了被试学习到的先验,发现该先验并非标准的均匀分布,而是在近似的均匀分布的两侧存在较长的尾巴;Chang 和 Jazayeri (2018)将各个时距的似然性与离散均匀分布进行卷积,构建的卷积先验对趋中效应行为数据的拟合度优于均匀先验;迭代贝叶斯估计模型则认为个体的先验为正态分布,其随着个体的学习过程而不断更新(Petzschner & Glasauer, 2011; Petzschner et al., 2015)。以上的研究在不同假设的基础上构建了时间信息的先验,均成功地预测了趋中效应。Zhang 和 Zhou(2017)采用时间复制任务,让被试按键复制随机混合呈现的视觉与听觉时距,发现视听时距信息形成了共同的时间先验的超通道表征(supramodal representation)。Roach等(2017)也在时间复制任务中证实了时间先验的超通道表征,并进一步发现消极呈现时距信息(不需要任何行为反应)、时距辨别和发音复制时距这三种任务类型均与按键复制时距任务不会形成共同的时间先验,提示感觉通道与任务类型会共同影响时间先验的形成。Zhang 和 Stocker(2022)发现,根据行为数据构建出的速度知觉的先验(行为先验),能够有效预测MT区域(视觉V5区)视觉速度群编码(population coding)的精确性(神经先验)。可见,通过将神经科学的技术与贝叶斯模型相结合,有望采用神经电生理证据检验以上研究中所提出的先验的合理性。
第二,厘清时间信息似然性的心理表征。有研究根据韦伯定律采用了正态分布似然性,也有研究根据费希纳定律认同对数正态分布似然性。正态分布与对数正态分布似然性均能很好地拟合趋中效应的行为数据(Jazayeri & Shadlen, 2010; Petzschner & Glasauer, 2011),但是对数正态分布似然性对Kappa效应行为数据的拟合度优于正态分布似然性(Chen et al., 2021)。Toso等(2021)通过记录大鼠神经元活动,证明背外侧纹状体只对客观时间而非主观时间进行编码。概率群编码认为贝叶斯推理建立在大脑神经群活动的基础上,可以通过测量输入信息诱发的神经群活动来解码该信息的似然性(Walker et al., 2020)。未来的研究可以进一步在背外侧纹状体测量客观时间编码相关的神经活动,对似然性进行解码,从而厘清时间信息似然性的心理表征是正态还是对数正态分布。
第三,探明时间信息贝叶斯估计的神经基础。Jazayeri 和 Shadlen(2015)发现猴子顶叶神经元的放电模式与趋中效应有关。在脑电研究方面,已有研究指出P3成分与贝叶斯推理有关(Kopp et al., 2016)。与这些研究一致,Cui等(2022)发现源于顶叶的P2和P3b成分与时空信息的贝叶斯整合有关。Hayashi 和 Ivry (2020)测量顶叶皮质的时距调谐曲线(tuning curve),证明顶叶皮质编码主观时间(后验)而非客观时间(似然性)。根据以上的研究,时间信息的贝叶斯估计可能发生于顶叶皮质。在计算模型方面,Pérez 和 Merchant(2018)构建了递归神经网络,Narain等(2018)根据小脑的解剖结构构建了小脑计算模型,分别模拟了人脑加工时间信息的贝叶斯推理过程,预测了时间知觉的趋中效应。这些研究说明时间信息的贝叶斯估计可能涉及更大范围的神经网络,并非是顶叶特异的。未来的研究需要进一步提供证据,探明时间信息的贝叶斯估计是顶叶特异还是涉及更大范围的神经网络。
时间信息加工模型和贝叶斯推理联系密切,也并非完全等同。大量的研究发现注意、唤醒和情绪能够调节时间知觉(Droit-Volet & Meck, 2007),如何将注意、唤醒和情绪因素纳入贝叶斯模型还有待探讨。一种方案是在标准贝叶斯模型中引入修正系数,例如,Chen等(2018)让被试注意视觉信息而忽略听觉刺激,发现视听时距信息的估计值是视觉和听觉时距信息的加权平均值,但其加权系数是标准贝叶斯模型预测的加权系数与一个修正系数的乘积。Yoshimatsu 和 Yotsumoto (2021)也报告通道特异的注意会对视听时间信息整合的加权系数进行修正。一种不同于修正系数的可能性是:注意、唤醒和情绪调节时间信息的似然性,然后标准贝叶斯模型将先验与调节后的似然性整合成为后验。时间信息加工模型对时间信息的各个加工阶段进行了较为详细的描述,注意、唤醒和情绪可以调节时钟阶段累加的脉冲数量(似然性),进而影响主观时间的心理表征(Droit-Volet & Meck, 2007)。可见,未来应该结合时间信息加工模型与贝叶斯模型的优势,为时间信息加工的认知神经机制研究提供新的思路。
参考文献
Acerbi, L., Wolpert, D. M., & Vijayakumar, S. (2012). Internal representations of temporal statistics and feedback calibrate motor-sensory interval timing. PLoS Computational Biology, 8(11), Article e1002771. https://doi.org/10.1371/journal.pcbi.1002771
Adams, W. J. (2016). The development of audio-visual integration for temporal judgements. PLoS Computational Biology, 12(4), Article e1004865. https://doi.org/10.1371/journal.pcbi.1004865
Billock, V. A., & Tsou, B. H. (2011). To honor Fechner and obey Stevens: Relationships between psychophysical and neural nonlinearities. Psychological Bulletin, 137(1), 1–18.
Brock, J. (2012). Alternative Bayesian accounts of autistic perception: Comment on Pellicano and Burr. Trends in Cognitive Sciences, 16(12), 573–574.
Burr, D., Della Rocca, E., & Morrone, M. C. (2013). Contextual effects in interval-duration judgements in vision, audition and touch. Experimental Brain Research, 230(1), 87–98.
Casassus, M., Poliakoff, E., Gowen, E., Poole, D., & Jones, L. A. (2019). Time perception and autistic spectrum condition: A systematic review. Autism Research, 12(10), 1440–1462.
Chang, C.-J., & Jazayeri, M. (2018). Integration of speed and time for estimating time to contact. Proceedings of the National Academy of Sciences, 115(12), E2879–E2887.
Chen, L. H., Zhou, X. L., Müller, H. J., & Shi, Z. H. (2018). What you see depends on what you hear: Temporal averaging and crossmodal integration. Journal of Experimental Psychology: General, 147(12), 1851–1864.
Chen, Y. G., Peng, C. H., & Avitt, A. (2021). A unifying Bayesian framework accounting for spatiotemporal interferences with a deceleration tendency. Vision Research, 187, 66–74.
Chen, Y. G., Zhang, B. W., & Körding, K. P. (2016). Speed constancy or only slowness: What drives the Kappa effect. PLoS ONE, 11(4), Article e0154013. https://doi.org/10.1371/journal.pone.0154013
Cicchini, G. M., Arrighi, R., Cecchetti, L., Giusti, M., & Burr, D. C. (2012). Optimal encoding of interval timing in expert percussionists. Journal of Neuroscience, 32(3), 1056–1060.
Cohen, J., Hansel, C. E. M., & Sylvester, J. D. (1953). A new phenomenon in time judgment. Nature, 172(4385), Article 901. https://doi.org/10.1038/172901a0
Cui, M. H., Peng, C. H., Huang, M., & Chen, Y. G. (2022). Electrophysiological evidence for a common magnitude representation of spatiotemporal information in working memory. Cerebral Cortex, 32(18), 4068–4079.
Droit-Volet, S., & Meck, W. H. (2007). How emotions colour our perception of time. Trends in Cognitive Sciences, 11(12), 504–513.
Elliott, M. T., Wing, A. M., & Welchman, A. E. (2014). Moving in time: Bayesian causal inference explains movement coordination to auditory beats. Proceedings of the Royal Society B: Biological Sciences, 281(1786), Article 20140751. https://doi.org/10.1098/rspb.2014.0751
Ernst, M. O., & Banks, M. S. (2002). Humans integrate visual and haptic information in a statistically optimal fashion. Nature, 415(6870), 429–433.
Gibbon, J. (1977). Scalar expectancy theory and Weber’s law in animal timing. Psychological Review, 84(3), 279–325.
Gori, M., Chilosi, A., Forli, F., & Burr, D. (2017). Audio-visual temporal perception in children with restored hearing. Neuropsychologia, 99, 350–359.
Griffiths, T. L., & Tenenbaum, J. B. (2011). Predicting the future as Bayesian inference: People combine prior knowledge with observations when estimating duration and extent. Journal of Experimental Psychology: General, 140(4), 725–743.
Gu, B. M., Jurkowski, A. J., Shi, Z. H., & Meck, W. H. (2016). Bayesian optimization of interval timing and biases in temporal memory as a function of temporal context, feedback, and dopamine levels in young, aged, and Parkinson’s disease patients. Timing & Time Perception, 4(4), 315–342.
Hallez, Q., Damsma, A., Rhodes, D., van Rijn, H., & Droit-Volet, S. (2019). The dynamic effect of context on interval timing in children and adults. Acta Psychologica, 192, 87–93.
Hayashi, M. J., & Ivry, R. B. (2020). Duration selectivity in right parietal cortex reflects the subjective experience of time. Journal of Neuroscience, 40(40), 7749–7758.
Helson, H. (1930). The tau effect—An example of psychological relativity. Science, 71(1847), 536–537.
Ide, M., Yaguchi, A., Sano, M., Fukatsu, R., & Wada, M. (2019). Higher tactile temporal resolution as a basis of hypersensitivity in individuals with autism spectrum disorder. Journal of Autism and Developmental Disorders, 49(1), 44–53.
Jazayeri, M., & Shadlen, M. N. (2010). Temporal context calibrates interval timing. Nature Neuroscience, 13(8), 1020–1026.
Jazayeri, M., & Shadlen, M. N. (2015). A neural mechanism for sensing and reproducing a time interval. Current Biology, 25(20), 2599–2609.
Jones, B., & Huang, Y. L. (1982). Space-time dependencies in psychophysical judgment of extent and duration: Algebraic models of the tau and kappa effects. Psychological Bulletin, 91(1), 128–142.
Karaminis, T., Cicchini, G. M., Neil, L., Cappagli, G., Aagten-Murphy, D., Burr, D., & Pellicano, E. (2016). Central tendency effects in time interval reproduction in autism. Scientific Reports, 6, Article 28570. https://doi.org/10.1038/srep28570
Kopp, B., Seer, C., Lange, F., Kluytmans, A., Kolossa, A., Fingscheidt, T., & Hoijtink, H. (2016). P300 amplitude variations, prior probabilities, and likelihoods: A Bayesian ERP study. Cognitive, Affective & Behavioral Neuroscience, 16(5), 911–928.
Körding, K. P., Beierholm, U., Ma, W. J., Quartz, S., Tenenbaum, J. B., & Shams, L. (2007). Causal inference in multisensory perception. PLoS ONE, 2(9), Article e943. https://doi.org/10.1371/journal.pone.0000943
Miyazaki, M., Yamamoto, S., Uchida, S., & Kitazawa, S. (2006). Bayesian calibration of simultaneity in tactile temporal order judgment. Nature Neuroscience, 9(7), 875–877.
Narain, D., Remington, E. D., Zeeuw, C. I. D., & Jazayeri, M. (2018). A cerebellar mechanism for learning prior distributions of time intervals. Nature Communications, 9(1), Article 469. https://doi.org/10.1038/s41467-017-02516-x
Pellicano, E., & Burr, D. (2012). When the world becomes “too real”: A Bayesian explanation of autistic perception. Trends in Cognitive Sciences, 16(10), 504–510.
Pérez, O., & Merchant, H. (2018). The synaptic properties of cells define the hallmarks of interval timing in a recurrent neural network. Journal of Neuroscience, 38(17), 4186–4199.
Petzschner, F. H., & Glasauer, S. (2011). Iterative Bayesian estimation as an explanation for range and regression effects: A study on human path integration. Journal of Neuroscience, 31(47), 17220–17229.
Petzschner, F. H., Glasauer, S., & Stephan, K. E. (2015). A Bayesian perspective on magnitude estimation. Trends in Cognitive Sciences, 19(5), 285–293.
Pöppel, E. (1997). A hierarchical model of temporal perception. Trends in Cognitive Sciences, 1(2), 56–61.
Pouget, A., Beck, J. M., Ma, W. J., & Latham, P. E. (2013). Probabilistic brains: Knowns and unknowns. Nature Neuroscience, 16(9), 1170–1178.
Rescorla, M. (2021). Bayesian modeling of the mind: From norms to neurons. WIREs Cognitive Science, 12(1), Article e1540. https://doi.org/10.1002/wcs.1540
Roach, N. W., McGraw, P. V., Whitaker, D. J., & Heron, J. (2017). Generalization of prior information for rapid Bayesian time estimation. Proceedings of the National Academy of Sciences of the United States of America, 114(2), 412–417.
Shams, L., & Beierholm, U. R. (2010). Causal inference in perception. Trends in Cognitive Sciences, 14(9), 425–432.
Shi, Z. H., Church, R. M., & Meck, W. H. (2013). Bayesian optimization of time perception. Trends in Cognitive Sciences, 17(11), 556–564.
Toso, A., Reinartz, S., Pulecchi, F., & Diamond, M. E. (2021). Time coding in rat dorsolateral striatum. Neuron, 109(22), 3663–3673.e6.
Wada, M., Umesawa, Y., Sano, M., Tajima, S., Kumagaya, S., & Miyazaki, M. (2023). Weakened Bayesian calibration for tactile temporal order judgment in individuals with higher autistic traits. Journal of Autism and Developmental Disorders, 53(1), 378–389.
Walker, E. Y., Cotton, R. J., Ma, W. J., & Tolias, A. S. (2020). A neural basis of probabilistic computation in visual cortex. Nature Neuroscience, 23(1), 122–129.
Wing, A. M., Doumas, M., & Welchman, A. E. (2010). Combining multisensory temporal information for movement synchronisation. Experimental Brain Research, 200(3–4), 277–282.
Yoshimatsu, H., & Yotsumoto, Y. (2021). Weighted integration of duration information across visual and auditory modality is influenced by modality-specific attention. Frontiers in Human Neuroscience, 15, Article 725449. https://doi.org/10.3389/fnhum.2021.725449
Zhang, H. H., & Zhou, X. L. (2017). Supramodal representation of temporal priors calibrates interval timing. Journal of Neurophysiology, 118(2), 1244–1256.
Zhang, L. Q., & Stocker, A. A. (2022). Prior expectations in visual speed perception predict encoding characteristics of neurons in area MT. Journal of Neuroscience, 42(14), 2951–2962.
Zhou, H. Y., Cai, X. L., Weigl, M., Bang, P., Cheung, E. F. C., & Chan, R. C. K. (2018). Multisensory temporal binding window in autism spectrum disorders and schizophrenia spectrum disorders: A systematic review and meta-analysis. Neuroscience & Biobehavioral Reviews, 86, 66–76.
Bayes-optimal Perception of Temporal Information
Chen Youguo1,2, Peng Chunhua3, Liu Peiduo1,2, Yu Jie1,2
(1Time Psychology Research Center, Faculty of Psychology, Southwest University, Chongqing, 400715)
(2Center of Studies for Psychology and Social Development, Southwest University, Chongqing, 400715)
(3Chongqing Key Laboratory of Emotion and Mental Health, Chongqing University of Arts and Sciences, Chongqing 402160)
Abstract:
The Bayesian theory is widely used in cognitive science. Previous studies have proven that Bayesian inferences cause many illusions in temporal cognition. From the perspective of Bayesian inference, we analyzed the processing mechanism of the central tendency effect, integration of multisource temporal information, spatiotemporal interference effect, and Bayesian calibration of simultaneity. This study has crucial implications for research on temporal information processing; additionally, it helps other researchers in cognitive science to understand the basic methods of modeling mental processes using the Bayesian theory.
Temporal information processing is reinterpreted using a Bayesian model. The clock, reference memory, working memory, and decision-making in the temporal information processing model were analogized with the likelihood, prior, posterior, and loss functions of Bayesian inference. The central tendency effect is known as the Vierordt’s law, in which participants underestimate long time intervals and overestimate short time intervals in various time intervals. A three-stage Bayesian model is constructed to explain the mechanism of the central tendency effect. In daily life, to the integration of multiple sources of temporal information is necessary. The maximum likelihood estimation model proposes that the brain uses Bayes’ rule to integrate temporal information from different sources, reducing uncertainty and increasing estimation reliability. The causal inference model uses a hierarchical Bayesian model to determine whether different temporal information originate from the same source or different sources. The spatiotemporal interference effect involves the mutual interference between spatial and temporal information, among which the Kappa effect has significantly progressed. The Kappa effect is a spatiotemporal illusion in which the irrelevant distance between the stimuli systematically distorts the perception of elapsed time between sensory stimuli. An algebraic model assumes that the perceived interstimulus time is a weighted average of actual and expected times, calculated as a ratio of known distance and velocity. This algebraic model was rewritten as a Bayesian model with a constant-speed hypothesis. A logarithmic constant-velocity model was proposed by integrating the Weber-Fechner law with an algebraic model. The logarithmic constant-velocity model considers that the deceleration tendency of the Kappa effect is driven by the Weber-Fechner law. The fitness of the logarithmic model for the Kappa effect behavior data is better than that of the original constant-velocity model. Additionally, priors influence the simultaneity of temporal order perception. During the temporal-order judgment task, participants learned the statistical distribution of temporal-order information as a prior. The prior and likelihood of the temporal-order information are then integrated using the Bayes’ rule, which is similar to the central tendency effect.
Future research on Bayes-optimal perception of temporal information should answer the following questions. (1) Test the rationality of the prior proposed in previous studies. Neuroscience should be combined with the Bayesian model to test priors using electrophysiological evidence. (2) Clarify the mental representation of the likelihood of temporal information. Neural activity in the dorsolateral striatum is associated with objective temporal coding. Neural activity can be measured to decode the likelihood of temporal information and to clarify whether the likelihood of temporal information has a normal or lognormal distribution. (3) Identify the neural basis of the Bayesian estimation of temporal information. Further evidence is needed to identify whether temporal Bayesian estimation is specific to the parietal cortex or involves a larger neural network. The advantages of the temporal information processing and Bayesian models should be combined to provide new ideas for research on the cognitive and neural mechanisms of temporal information processing.
Keywords: temporal information, Bayesian inference, central tendency effect, temporal information integration, spatiotemporal interference effect, Bayesian calibration of simultaneity
https://m.sciencenet.cn/blog-3400702-1420300.html
上一篇:且思且行,致远致精——记参加第二十五届全国心理学学术会议
下一篇:文献报告:社会拥挤对自我提升类产品偏好的影响