博文
分子系统学对有刺蜂类进化以及蚂蚁和蜜蜂起源的观点
 精选
精选
||||
分子系统学对有刺蜂类进化以及蚂蚁和蜜蜂起源的观点
Branstetter et al., 2017, Current Biology 27, 1019–1025April 3, 2017 Published by Elsevier Ltd.http://dx.doi.org/10.1016/j.cub.2017.03.027
Phylogenomic Insights into the Evolution of Stinging Wasps and the Origins of Ants and Bees
翻译:中国科学院动物研究所博士后,李晓莉博士
原文:
Phylogenomic Insights into the Evolution of Stinging Wasps and the Origins of Ants.pdf
亮点
1、UCE系统发育极好解决了有刺蜂类系统发育问题。
2、蚂蚁是采花酿蜜的蜜蜂和有刺蜂类的姐妹群。
3、蜜蜂嵌套在方头泥蜂科中,是短柄泥蜂亚科和大头泥蜂亚科的姐妹群。
4、外群的选择和分类取样极大影响分子系统学的结果。
总结
有刺蜂类(膜翅目:针尾部)是膜翅目昆虫中极具多样性的类群。目前,已经涵盖超过70000种描述和多样性的生活史记载,包括体外寄生,盗食寄生,捕食性以及取食花粉的蜂类(蜜蜂 [喜花类]和棒角蜂科)和真社会性昆虫(群居的胡蜂、蚂蚁和一些蜜蜂)[1]。目前对谱系关系研究最好的是在大多数陆地生态系统中占有生态优势的蚂蚁[2],蜜蜂以及被子植物主要的传粉昆虫[3]。构建蚂蚁和蜜蜂的进化亲缘关系有助于我们理解和重建社会进化模式以及充分鉴别从肉食性向取食花粉的食性改变的生物学意义。尽管最近针尾部的亲缘关系研究[4–11]取得了一定的进展,但是在针尾部高级阶元的亲缘关系中仍然存在相当多的不确定性,如蚂蚁和蜜蜂的亲缘关系[5–7]。我们用超级保守元件(UCE)系统基因组学方法[7,12] ,利用> 800 UCE位点和从187头标本采集到的序列数据,包括30-31个针尾部的科的数据,解决有刺蜂类科级阶元之间的关系。我们用多种分析方法对187个分类单元数据集进行了分析,并对几种分类单元数据集进行了分析评价。我们还测试了蚂蚁和蜜蜂系统发育位置的替代假说。我们得出的结论高度支持了有刺蜂类的系统发育关系。最重要的是,我们发现了明确的证据表明:蚂蚁和蜜蜂以及有刺蜂(蜜蜂总科)是姊妹群;蜜蜂是作为并系群嵌套在方头泥蜂科中。我们还证明了分类单元的选择可以从根本上影响系统发育树的拓扑结构和在分子系统学推理的结果。
结果与讨论
分子系统学的分析
我们采用分子系统学方法并结合超级保守元件靶向富集方法(UCE)的新一代测序(NGS)技术,解决主要有刺蜂类分支之间的关系(总科和科)[12]。UCE的方法依赖于DNA和从新鲜标本以及博物馆保存的标本中提取的数百个位点的有效排序。我们随后发表了实验室报告[7,12](参见所附实验流程)和使用膜翅目特有引物富集1510个位点 [7]。我们从139个分类单元中测序得出新的分子数据,将这些数据与先前得到的16个种测序的结果[7]以及在UCE位点的得到的32个基因组数据放在一起,最终形成包含187个分类单元的数据集(见数据S1样品信息)。
我们的研究包括136个有刺蜂类样本,有30-31个常见科,只缺少菱板蜂科。我们对有刺蜂蜂和蜜蜂(蜜蜂总科)进行密集取样,包括23-25个亚科共计53种,并选取外群16种的蜜蜂,包括系统发育种较为少见的家族长背泥蜂科和Heterogynaidae。我们选取了方头泥蜂科4-8个亚科14种的标本,其中包括2个假设与蜜蜂密切相关的亚科(短柄泥蜂亚科和大头泥蜂亚科)[5]。前人研究提出了与针尾部密切相关的类群[8, 10, 13–15]。因此,我们选取叶蜂(广腰亚目)和8-12个细腰亚目(寄生部)非蛰刺蜂总科的标本,包括钩腹蜂总科,旗腹蜂总科,姬蜂总科和分盾细蜂总科作为外群。。
富集的UCE的位点,我们平均从每个类群捕获测序了966个片段。其中,有801bp平均序列长度和平均每校每803个重叠覆盖率(见数据S1装配信息)。我们评估、筛选并比较不同级别的分类单元的影响百分比(在给定的时间内所需的分类数百分比位点),最后选定75%的筛选比例(''hym-187t-f75”)作为分析的主要数据集。此数据集包括854序列数据的位点以及203095 bp的序列长度,其中包含143608个位点的信息(见对齐矩阵信息的数据S1)。我们采用最大似然法(MLhym-187t-f75数据集;RAxML V8[16])、贝叶斯推断(BI;exabayes v1.4 [17]),和物种树分析(ST;astral-ii [18])的方法。对于ML分析,我们比较了几种不同的数据分区方案(见更多信息的数据S1)和为减轻碱基组成异质性或替换饱和引起的系统发育误差的两种方法。对于后者的方法,我们创建了一个转换的数据集,整个矩阵到非编码,其中一个我们去掉了显示碱基组成异质性的位点的分类群(除去47个位点)。对于ST分析,我们采用加权统计分级减少低信息位点错误内容[19]。
我们重建了一个支持度较高的针尾部的系统发育树。所有的分析中拓扑结构几乎相同(图1,S1,和S2)。我们观察到了8个节点的拓扑冲突,关系中最重要的区别的科是青蜂总科(cuckoowasps及其亲属)。我们发现钩腹蜂科作为有刺蜂的姊妹群(针尾部)在所有分析中都得到最大支持。虽然在我们的数据集种缺乏几个胡蜂总科的数据,但这个结果是与最近的分子分析一致[ 8, 10,15 ]。重要的是,我们在任何分析中没有得到姬蜂总科与有刺蜂有密切关系的结论。然而,姬蜂总科长期被作为针尾部的可能的姐妹群[13]。在针尾部中,我们发现青蜂科是一个并系群,其分支包含短节蜂科+ [梨头蜂科+螯蜂科]。在大多数系统发育分析中,将其作为姐妹群重建到其它非青蜂科中。在所有分析中,其余的针尾部的总科都有两个主要分支,均得到高度支持。总的来说,总科之间的关系很大程度上支持最近的转录组分析结果[6],除了对胡蜂总科的定位。除了编码的ML分析(96%)和ST分析(90%),在所有的分析中,蚂蚁(蚁科)被推断为蜜蜂和有刺蜂的姐妹群。
对于包含在蜜蜂和有刺蜂中(蜜蜂总科)的分支,我们对科及以上阶元关系的研究结果表明分支在分析中是相同的。这基本符合Debevec等学者同意的分支[5]。最重要的是,我们发现蜜蜂(Anthophila)被嵌套在并系群方头泥蜂科和姊妹群短柄泥蜂科+大头泥蜂亚科中。这个发现首次被4个分子标记的研究工作中报道[5]。但当时的数据只适度的支持。在蜜蜂总科,我们还发现长背泥蜂科是有刺蜂和Heterogynaidae的姊妹群。Heterogynaidae是一个非常神秘的科,是方头泥蜂亚科+泥蜂科的姐妹群。在蜜蜂(Anthophila),我们对于高级阶元关系的研究结果与以前的基本上是一致的[20]。最值得注意的是,我们发现Melittidae是蜜蜂科的姐妹群,我们重建了真社会性后足具花粉筐的蜂类(蜜蜂族、Bombini和Meliponini)单系群的地位,并把所有的联合起来进行分析(未恢复ST分析)。
检验蚂蚁和蜜蜂的系统发育关系
在界定蚂蚁和蜜蜂的系统发育位置时,有刺蜂是至关重要的。蚂蚁是在几乎所有陆地生态系统中占主导地位的社会性昆虫,而蜜蜂是世界上最大的维管束植物—被子植物最重要的传粉者之一。为了评估我们的系统发育结果的稳定性,我们开展了两类分析。首先,我们使用ShimodairaHasegawa(SH)评估了一系列先前提出的蚂蚁和蜜蜂的系统发育假说 [21]。对于SH测试,考虑到先前两个类群的系统发育假说(蚂蚁[4, 6, 8,9];蜜蜂[5, 22 - 25]),我们分析了蚂蚁的九个交替位点和蜜蜂的14个交替位点(表1)。我们用最丰富的187种分类数据集进行了测试,其中包含100种类群较为平衡的数据集。在所有情况下,替代拓扑被拒绝(p<0.01),为这里所介绍的首选拓扑提供了明确的支持。根据这些分析和我国目前的分类取样水平来看,蚂蚁是蜜蜂总科的姐妹群,蜜蜂显然是方头泥蜂科的高度衍生类群。
其次,我们分析了分类单元取样对系统发育结果的影响。先前的分子系统学研究中所包括的分类群比我们得到的蚂蚁与蜜蜂总科的定位冲突时使用的分类群少得多。Johnson等人基于转录组的研究[6]发现蚂蚁是蜜蜂总科的姐妹群(这里的结果)。鉴于Faircloth等人对于UCE的基础研究[7],发现蚂蚁同样是其它大部分针尾部的姐妹群,而青蜂总科并不包括在他们的分析中。我们将分类单元取样试验分为以下几类(图2):(1)Johnson等人的数据[6],(2Faircloth等人的数据[7]和(3)当前分类数据的变化。在第一类中,产生了两个数据集,一个完全相同的分类片段为[6](''johnson-19t”)和一个与青蜂科argochrysisarmilla无关的片段(''johnson-18t”)。这个特别的操作是因为[ 6 ]和[ 7 ]之间的主要区别青蜂总科是否是其他所有针尾部类群的姐妹群。对于Faircloth等人的数据[ 7 ],我们重新创建原始的45种数据矩阵(''faircloth-45t”)和几种不同的分类集。首先,我们添加一个青蜂科(''faircloth-46t”),然后我们继续添加额外的针尾部平衡各主要支系(''faircloth-52t,' 'faircloth-56t,”和“'faircloth-61t”)。我们还通过从原始数据集中删除过量的蚂蚁类群数据来平衡数据集(''faircloth-26t”),然后添加一个青蜂科(''faircloth-27t”)。对于第三类,我们生成的数据集是将大部分外群删除(''hym-147t”),将金小蜂作为最先分开的外群,将大痣细蜂科(分盾细蜂总科),旗腹蜂总科,钩腹蜂总科作为最近的外群。在这个分类矩阵中,我们删除了青蜂(''hym-131t”)以及青蜂和钩腹蜂(‘‘Hym-131T’’)。同时,在经过深思熟虑之后删除过量的蚂蚁、蜜蜂和黄蜂类群的类群数据以平衡数据矩阵(''hym-100t”)。
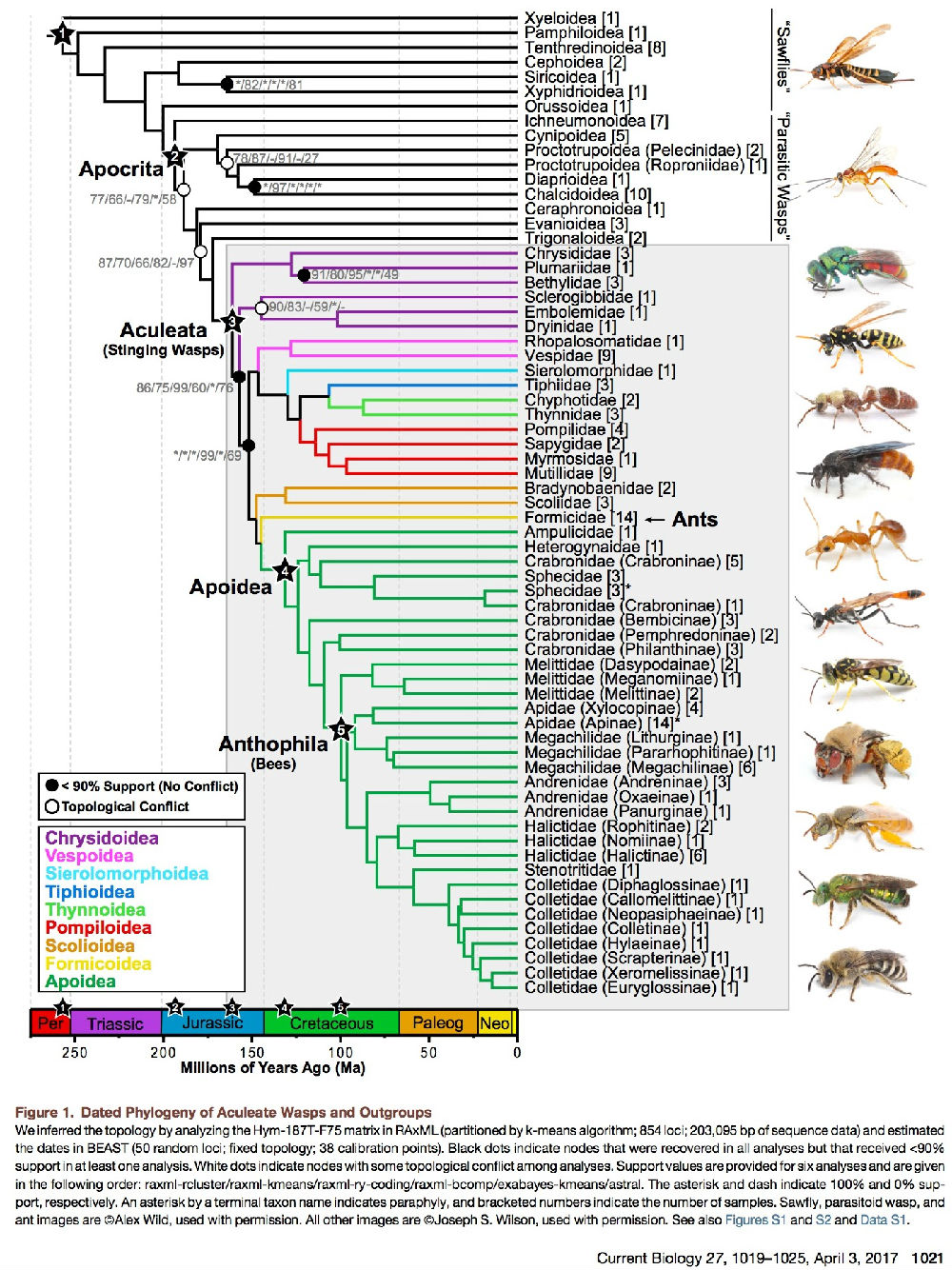
图1. 先前的针尾部蜂类和外群的系统发育关系
我们根据拓扑的RAxML和hym-187t-f75矩阵分析(k-均值算法;划分854个位点;203095 bp的序列数据)和估算BEAST的年代(50个随机位点;固定的拓扑结构;38个校准点)。黑点表示在所有分析中都恢复的节点,得到的结果< 90%,至少支持一种分析。白色圆点是节点之间的拓扑冲突分析。提供了六个分解的有价值的值,按以下顺序给出:raxml-rcluster/raxml-kmeans/raxml-ry-coding/raxml-bcomp/exabayes-kmeans/astral.
星号和破折号分别表示100%和0%支持。由终端分类单元的名称为星号表示,括号内的数字表示样本数。叶蜂,寄生蜂,和蚂蚁的图片已经得到Alex Wild的许可。所有其他的图像已获得Joseph S. Wilson的许可。参见图S1和S2和S1的数据。
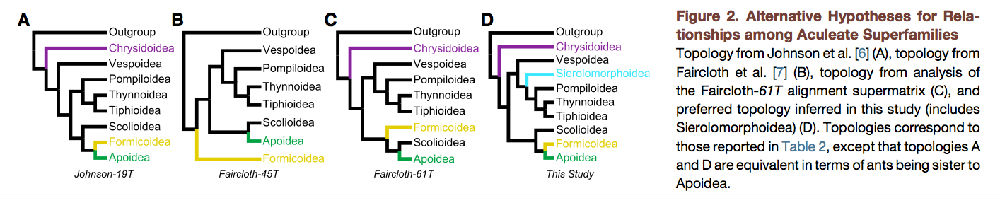
图2. 针尾部总科亲缘关系的替代假说
约翰逊等人的拓扑结构 [ 6 ](A),费尔克洛思等的拓扑结构[ 7 ](B),对faircloth-61t准超矩阵的拓扑分析(C),在这项研究中推断出的首选拓扑结构(包括瘤角蜂科)(D)。拓扑对应表2中所列的,拓扑A和D表明蚂蚁是蜜蜂总科的姐妹群,且进化的地位是相当的。

我们使用完整的187分类矩阵和平衡100种分类矩阵进行了非分区分析,我们支持的拓扑结构在所有情况下都明显优于替代方案。D(LH),似然分数差;alt.:交变、替代;excl.:除去,排除;incl.:包括;NA不适用。

结果表明,外群的选择(青蜂科存在/不存在)和类群均匀度可以影响结果。矩阵名称表示分类单元集是否是约翰逊等人[ 6 ]或费尔克洛思等人[ 7 ],或本研究(膜翅目)。恢复三种不同的拓扑结构:蚂蚁是蜜蜂总科的姐妹群(A);蚂蚁是其他所有针尾部总科的姐妹群,青蜂科除外(B);蚂蚁是蜜蜂总科+ 土蜂总科的姐妹群(C)。自举支持表示支持分支,包括蚂蚁加上它的姐妹群。拓扑与图1A - 1C中所示的相对应,仅与蚂蚁的位置相关。BS,引导支持。
分类单元取样试验的结果(表2)支持的结论是,即使是基因组规模的数据,外群的选择和分类的平衡都影响系统进化结果。Faircloth等人[7]的研究同样受这两个问题的影响。我们怀疑其进化树的研究中蚂蚁的定位研究是错误的。聚焦蚂蚁的位置(蚁总科:蚁科),我们重建了三种替代拓扑结构(图2;表2):蚂蚁是蜜蜂总科的姐妹群(拓扑A);蚂蚁是其他所有类群的姐妹群,青蜂总科除外(拓扑B);蚂蚁是蜜蜂总科和土蜂总科的姐妹群(拓扑C)。在Johnson等人的[6]矩阵,我们重建了拓扑A。分析先前Faircloth等的研究中[ 7 ]分类位置(faircloth-45t)产生的拓扑结构B。对于Faircloth-46t,Faircloth-52t,和Faircloth-56t,我们也重建了拓扑结构,然而,在Faircloth-61t分析、拓扑转移到C,将蚂蚁作为土蜂总科和蜜蜂总科的姐妹群。Faircloth-56t与Faircloth-61t之间的差异是增加了几个青蜂总科(梨头蜂科和螯蜂科),蛐蜂科(胡蜂总科)和长背泥蜂科(蜜蜂总科),后面两个类群打破了长分支。减少和平衡Faircloth-45t分类群也改变了相应的拓扑。减少蚂蚁的数量从22Faircloth-45t分类群到三种分类Faircloth-26t改变拓扑到A。Hym-147t矩阵和变异(hym-133t,hym-131t,和hym-100t)也产生了拓扑A。对于Hym-100t矩阵,我们减少了蚂蚁和蜜蜂,以平衡较大的分类设置,所有的关系,除了蚂蚁的位置外,ML和Hym-187t的矩阵分析都是相同的。除了拓扑的差异只是描述,从矩阵除去外群(青蜂科或钩腹蜂科)通常导致蚂蚁位置引导分数下降(表2)
生物学含义
我们的研究结果解决了长期存在的关于针尾部系统发育关系的争论,有助于进一步理解主要的传粉蜜蜂多样性以及主要的真社会性昆虫蚂蚁的多样性和生态优势。蚂蚁与蜜蜂具有惊人的紧密关系,从而影响我们如何看待其重要行为的进化,如筑巢,中心地点觅食和针尾部蜂类的群居[6]。重要的是要强调以下事实:真社会性蚂蚁一次的进化,至少是蜜蜂进化的6-8倍[20]。这意味着包含蚂蚁和蜜蜂的分支可能是特别事先准备好进化为群居昆虫。如[6]所述,了解蜜蜂总科(蜜蜂和有刺蜂)所有的支系内的生物学,对深入了解生物学因素促进社会行为的进化具有重要意义。
我们的研究结果基本证实了先前的研究结果对于蜜蜂总科[5]和蜜蜂[20]的亲缘关系。我们确认将长背泥蜂科作为蜜蜂总科剩余的姐妹群,将蜜蜂作为方头泥蜂科下大头泥蜂亚科和短柄泥蜂亚科的姐妹群。蜜蜂与方头泥蜂科下大头泥蜂亚科和短柄泥蜂亚科的亲密关系以前在两项研究中都曾提出过形态学和分子数据(回顾[5])。大头泥蜂亚科包括在地面筑巢的蜂类,捕食各种各样的猎物,包括甲虫,蚂蚁,甚至蜜蜂。短柄泥蜂亚科包括小型,大多是洞巢蜂类,捕猎多样化,包括弹尾目(跳虫),缨尾目(蓟马)以及一些植食性半翅目类群(蚜虫,蚧壳虫,木虱,叶蝉, 诛蝉和角蝉)。短柄泥蜂亚科和大头泥蜂亚科合在一起,包含了超过2200个种类[26]。蜜蜂已经有20000余种,仅仅跟2200种捕食性蜂类是姊妹群。这暗示了这种从捕食到传粉的食性的转换是蜜蜂多样性的重要驱动力。未来的研究应包括更广泛的采样和测试大头泥蜂亚科和短柄泥蜂亚科这一假说。
先前基于形态学认为准蜂科[27]是长舌蜜蜂的姐妹群(蜜蜂科+切叶蜂科)。我们在蜜蜂的研究中进一步证实了准蜂科是一个单系群,为其它蜜蜂所有科的姐妹群。蜜蜂的科及关系与先前的研究完全一致[20]。值得注意的是,我们的大多数分析重建发现真社会性具花粉筐的蜜蜂(蜜蜂,大黄蜂,和无刺蜂)作为单系群和弱社会性昆虫Euglossini(orchid bees)的姐妹群,从而有利于组内的群居性的一个单一的起源。这些类群之间的关系一直存在争议,但我们结果与最近的分子系统学研究发现,控制碱基组成的异质性有利于真社会性具花粉筐昆虫保持单系性[28]。
结论
减少代表类群取样,同时开展NGS将驱动了分子系统学革命,使它可以用传统方法的一小部分成本生成较大的数据集[ 29, 30 ]。在这里,我们进一步把UCEs[ 12 ]靶向富集这样一个有前途的方法,成功应用于膜翅目等生物多样性丰富的昆虫。这极大地拓展了以前的研究,也是首次对肢动物使用的UCE的方法[ 7 ]。我们关注的是有刺蜂(针尾部)科级阶元的关系,同时产生一个强大的主干系统发育关系,并提供了许多关于这一进化历史的见解。此外,通过开展一系列的类群取样实验,我们已经证明,即使在系统发育基因组学时代,密集的分类取样是至关重要的,与外群的选择和分类的均匀性均会显著影响拓扑结构和自引支持率。
https://m.sciencenet.cn/blog-536560-1071872.html
上一篇:第十五届“中国昆虫学会分类与区系学术会议”会议总结
下一篇:周五学术报告(20170825)
全部作者的精选博文
全部作者的其他最新博文
全部精选博文导读
相关博文
- • Minerals线下恳谈会:履践致远、与时偕行——对话中国科学院广州地球化学研究所期刊合作学者
- • 聚英才 建高地 | 北京理工大学“特立青年学者”全球招聘开启
- • 700年后日本或濒临灭绝?日本学者推算预测:届时或仅剩1名15岁以下孩子
- • [转载]【同位素视角】非英语母语学者如何区分’e.g.’, ‘i.e.’, ‘namely’与‘such as’等混淆难题
- • 美国佐治亚大学等机构学者:刈割策略对Bulldog 805紫花苜蓿+Tifton 85狗牙根混播草地产量及品质的影响
- • 美国堪萨斯州立大学、密苏里大学等机构学者研究成果:土壤水分管理策略和品种多样性对紫花苜蓿产量、营养品质和农场盈利能力的影