博文
Journal Club:昆虫性别丢失:无性的原因和对基因组的影响(Tvedte等,2018)
 精选
精选
|||
昆虫性别丢失:无性的原因和对基因组的影响
Eric S Tvedte1 John M Logsdon Jr.2 Andrew A Forbes2
https://doi.org/10.1016/j.cois.2018.11.007
重点
自然种群中为什么保持性别仍然是进化 生物学中的一个基本问题。已有并验证了几个假说。
尽管昆虫生殖系统多种多样,但对有性系与无性系的比较研究却很少。
新一代测序数据集可以对性别丢失原因和后果进行基因组水平研究。
昆虫具有惊人的繁殖策略多样性,为过渡到无性生殖的原因和后果的比较研究提供了有吸引力的模型。 我们概述了昆虫生殖系统的一些当代研究,并建立了无性昆虫基因组资源的初始数据库。 在昆虫系统中,失去性别的各种机制已经产生了一些重要的见解,包括遗传、内共生介导和杂交。 性别丢失后的突变和替代研究为无性的假设效应提供了最强有力的经验支持,而生态假说则有一些混合证据,如无性系中寄生虫负载增加和生态位宽度改变。 大多数假设已经在少数几个类群(例如竹节虫、蚜虫)中进行了探索,而大部分昆虫的分类广泛性仍然未得到充分研究。 鉴于昆虫无性近因的变化,我们主张扩大研究系统的分类学范围。 尽管在调查昆虫的性别方面存在一些挑战,但基因组测序的成本效益提高使得生成近缘的无性和有性支系数据变得越来越可行。
介绍
“我们甚至一点不了解性的最终原因; 为什么新生物应该由两性结合产生,而不是通过一个过程孤雌生殖产生 。“
-查尔斯·达尔文[ 1 ]
生物体繁殖方式是其生物学的一个重要特征。 在动物中,有性生殖是主要的策略,但我们还没有全面理解为什么它比其它繁殖方式更受青睐。对有性支系近缘的无性支系的研究,有希望成为确定性别的益处和/或成本的途径。 昆虫的 繁殖 策略之间有许多过渡。它们是研究无性生殖起源和结果且特别容易处理的生物。 我们首先简要介绍一下自然界维持性别的主要假说。 然后,我们回顾了昆虫生殖系统多样性以及一些研究揭示生殖转变的后果。 最后,我们确定了无性昆虫研究的挑战,但提出:特别关注这些系统将有望在理解性别进化方面取得实质性进展。
为什么有性别?
性别的进化悖论一直是几个严格理论研究的主题。 虽然性别无处不在,但它与预期对适合度有害的明显成本(例如代谢、遗传、产生雄性)有关[ 2 , 3 , 4 , 5 , 6 ]。 在其它条件相同的情况下,无性生殖应该更有益- 至少在短期内- 无性支系应该胜过共存的有性支系。
没有普遍的解释,可以说明性的进化和维持。 然而,有个引发重组的假说:在有性个体中可以打破连锁基因座之间的干扰,并使选择能够作用于单个基因座而不依赖于它的 遗传背景 [ 7 , 8 ]。这可以在有性种群中快速汇集适应性等位基因[ 9 , 10 ]或防止有害等位基因的积累[ 11 ]。 相关的生态假说包括有性生物产生 遗传多样性, 作为逃避 寄生的手段 [ 12 , 13 ]或利用资源有限的异构环境[ 14 ]。 实际上,多种机制可以“多元方式”共同维持性别[ 15 , 16 ]。 选择生活史性状(例如 越冬 卵)可间接促进性别的持续存在[ 17 ];在这些情况下,研究人员应审慎地将适合度结果归因于生殖模式。
昆虫的繁殖转变
虽然昆虫中有性繁殖的普遍性反映了其它动物的百分比(即> 99%的物种有性生活),但它们的生殖模式多样性超过二元的“性”或“无性”( 方框1 )。 无性繁殖 精确模式在连续几代中决定了杂合的基因组模式 ,这可能会影响支系的进化轨迹[ 18 , 19 ]。 有关昆虫生殖转变 的综述 ,我们鼓励读者参考Vershinina&Kuznetsova最近的两部分评论[ 20 ]和Gokhman&Kuznetsova [21 ]。
方框1、昆虫无性繁殖的模式
专性孤雌生殖 (Obligate parthenogenesis):没有受精的胚胎生长和发育
兼性孤雌生殖 (Facultative parthenogenesis):个体之间无性繁殖和有性繁殖切换 ,通常是对环境压力的响应
循环孤雌生殖 (Cyclical parthenogenesis):几代无性繁殖后是有性生殖;经常季节性。
偶发孤雌生殖 (Tychoparthenogenesis):一小部分未受精卵自发孵化。
无融合生殖(Apomictic reproduction / Apomixis) :后代的克隆生产,雌性子代在遗传上几乎与母本相同。
自体融合(Automictic reproduction / Automixis) :减数分裂卵的生产,具有允许恢复亲本倍性的各种机制。
雌核发育(Gynogenesis) :用精子和卵子开始繁殖,然后立即丢弃父本基因组。
杂交发生 (Hybridogenesis):用精子和卵子开始繁殖,然后在一代后丢弃亲本基因组
Normark [ 22 ]提供了一个专性无性昆虫的系统发育分布。 无性繁殖转变在几个昆虫类群中很丰富,包括 蜉蝣( Ephemeroptera)、竹节虫(Phasmotodea)(棒状昆虫)和Paraneoptera(虱子、 蓟马 和真正的半翅目昆虫)。 大多数无性昆虫支系寿命短,使它们的系统发育分布呈现“萎缩”的形态[ 14 , 23 ]。 虽然这一观察结果支持无性化不会导致长期进化成功的观念,但另一种解释包括无性支系的不常见起源和/或回归有性生殖[ 22 , [24 ],虽然后者可能很少见。
昆虫无性的起源
有性祖先如何产生无性支系是一个基本问题,虽然它在很大程度上仍然没有答案,但昆虫中已经记录了多种机制。 首先,基因突变可以直接诱导生殖方式过渡。记录这类例子通常涉及一个或几个主要位点的影响,包括豌豆蚜虫 Acyrthosiphon pisum [ 25 ]、 寄生蜂 Lysiphlebus fabarum [ 26 ]和开普蜜蜂 Apis mellifera capensis [ 27 ]。 然而,遗传标记 和无性关联研究通常没有明确识别生殖转变的基因和遗传机制。 一个例外是 A.m. capensis ,它有个遗传区域与 单性生殖密切关联,并和D. melanogaster 与减数分裂 纺锤体-F蛋白CG12114相互作用转录因子同源[ 28 ]。 A.m. capensis 这个基因可变剪切,将促成一个伪蜂王表型,并打开诱导孤雌生殖生产雌性生产开关[ 29 ]。 非编码区域也可以促进生殖模式转换。 上述豌豆蚜虫 A. pisum,miRNA和piRNA的差异表达与生殖型转换相关,但还没有直接功能关联验证[ 30 , 31 ]。 无性突变基础已经涉及到各种其它昆虫系统,尽管其确切的机制尚不清楚[ 32 ],并且很难将导因与性别丢失的后果分开。
第二,内共生菌可以操纵昆虫的生殖系统,导致它们失去性。 Stouthamer等 首次描述了细菌和昆虫宿主之间的关系 [ 33 ],并用抗生素处理无性 Trichogramma,使它们恢复有性生殖。 从那以后,许多无性昆虫系统中已经描述了母系传递的细菌,例如[ 34 , 35 , 36 , 37 ]。内共生菌介导的无性生殖似乎在 膜翅目中特别普遍 [ 38 • ]。 在内共生菌和生殖模式之间建立关联具有挑战性,但是在确认的例子中可以探索无性的基因组结果。 大多数内共生菌介导的转变伴随着一种自体融合形式,导致完整纯合子后代,被称为的 配子复制[38 • ]。 这种生殖模式的无性系可能会暴露于高遗传负荷,这应该不利于长期的支系持续。
第三,有性亲本支系之间的杂交事件后可能会出现无性生殖。 研究认为竹节虫[ 39 , 40 ]、甲虫[ 41 , 42 ]、 蚱蜢 [ 43 ]和蚜虫 [ 44 , 45 ]无性生殖就是杂交起源。 杂交支系预计会显示出大量的基因组杂合度 [ 46 ]。 在Timema 属竹节虫中,新一代测序(NGS)数据用于证明杂种无性系与父系物种的等位基因多样性,并确定了父系变异是无性系中一个子集的[ 47 • ]。
昆虫无性的后果
NGS数据越来越多,研究人员可以在候选基因的小数据集和大型全基因组数据集中评估性别丢失的影响( 表1 )。 中性进化过程预计会降低一般的有性结构,包括生殖器和交配行为机制 ,潜在的基因应该反映这些变化。 在某些情况下,如果编码这些结构和功能的基因活性成为不必要的代谢负担,则可能会对其进行强烈的选择。 实际上,这些影响可能在性别之间不同,导致雄性特异性状的丢失(例如发现配偶、求爱 、精子生产)和/或雌性性状(如招引信号、 如信息素、色素沉着和听觉信号)[ 48 ]。 寄生蜂 Leptopilina claves 无性基因组包括16个在无性系中具有有害SNP的基因,其中15个在其它昆虫中具有雄性特异表达,而在雌性受精囊中表达了一个[ 49 •• ]。此外,由于所有无性系都不具有相同变异。这表明这些突变在现在分开的无性系中独立起源。 Timema 属中,在多个单独到无性的过渡过程中观察到精子储存和色素沉着中涉及的基因衰变[ 50 ]。
表1 、 使用昆虫系统调查性行为的假设。
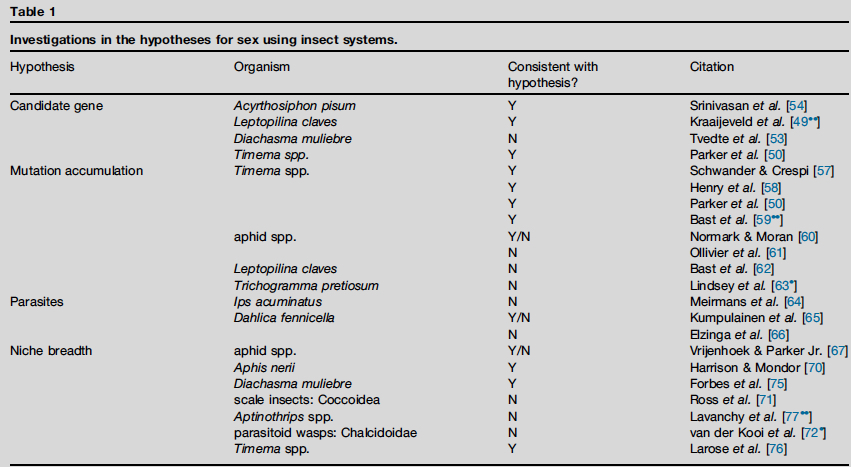
虽然无性基因组中性别特异性基因是退化靶标,但两性中有性生殖相关的基因也可能经历选择压力而变化。 例子包括减数分裂基因,它是有性繁殖所必需的,并在邻近真核生物起源事件进化[51 , 52 ]。 这些基因可能会经历依赖于无性繁殖精确模式的差异选择性方案。 在Diachasma 属寄生蜂有性和无性物种中,规范的减数分裂基因组件相同,表明无性繁殖通过自体融合[ 53 ]。 相比之下,豌豆蚜虫A. pisum基因组中几种减数分裂调控基因 发生了复制 [ 54 ]。 基因家族的扩张是 A. pisum 基因组的一般特征;参与胚胎发育 、发育和性别决定的基因中的重复事件, 尽管在减数分裂繁殖期减少,但 可以促进 生殖可塑性和维护遗传多样性 [ 55 ]。 减数分裂基因如何在无性克隆无性昆虫中进化,仍然是一个悬而未决的问题,但已经成熟用于未来的研究。 Timema 中,减数分裂纺锤体形成和中心体 组织途径过程中,多个生殖过渡均表现了基因表达差异[ 50 ]。
生殖模式与突变积累之间的关联研究结果好坏参半[ 56 • ]。Timema 属,无性竹节虫相较于有性物种,核和线粒体基因位点的小数据集显示较高突变负荷[ 57 , 58 ];Timema 属无性竹节虫转录组数据显示同义和非同义替换比例提升[ 59 •• ]。 然而,繁殖模式显然与豌豆蚜虫中的突变积累无关。 核基因EF1α在一个无性系中表现出加速的突变积累,但第二个支系缺乏这种模式,并且两个支系在COX2线粒体基因位点上均未显示任何清晰的模式[ 60 ]。 进一步,A. pisum 比较完整基因组和其它蚜虫表达序列标签(EST)的大数据集显示:有性和无性蚜虫支系的总体突变率和非同义替代率没有差异,同义率和dN / dS值的差异似乎是少数基因变化的结果[ 61 ]。 两个寄生蜂系统中,有性和无性基因组转座元件的累积类似[ 62 ,63 ]。 尽管分类资源丰富,但只有少数昆虫系统被用于研究生殖策略和生态变量之间的关系。为了证明生殖模式和寄生压力之间的关联,以前的研究已经使用共存有性和无性种群来度量a)种群中较高比例的有性个体 是否与更高寄生负荷一致,和b)假定无性种群的总体低遗传多样性是否会增加寄生感染的实际风险。 蠹虫 Ips acuminatus 种群中有害的寄生线虫流行与有性生殖个体比例无关[ 64 ]。 虽然在一项研究中发现了蓑蛾存在一种关联 [ 65 ],随后的一项研究表明:孤雌生殖胡蜂并不比同域分布的有性胡蜂更加容易被寄生。这可能是因为有性和无性物种之间羽化差异[ 66]。 据我们所知,尚未测量有性与无性昆虫在寄生压力下的遗传多样性。
在有性生殖丢失后,支系可用生态位可能会发生变化[ 67 ]。 对无性生殖物种可塑性的强烈选择可能有利于利用广泛生态位的,被称为“通用基因型”(GPG)假说[ 68 , 69 ]。 在蚜虫中,观测和系谱地理学证据通常支持少数优势显性基因型,从而产生单性生殖“超级克隆”[ 67 , 70 ,还见 67 例外]。 介壳虫 [ 71 ]和 小蜂 [ 72 • ] 宏分析也发现无性种群也比有性种群有更宽的生态位。 或者,过渡到无性生殖可能伴随其有性祖先的“冻结”遗传变异克隆 的快速生态位分割,这是“冻结生态变异”(FNV)假设的基础[ 73 , 74 ]。 各主要昆虫目支持FNV模型[ 67 , 75 ]。 但是,GPG和FNV模型并不相互排斥。 最近Timema中提出的一个潜在的情形,年轻和老年无性支系分别利用狭窄和宽生态位[ 76 ]。 同样重要的是在自然条件下验证实验结果。 例如,草蓟马的围栏实验 表明有性繁殖的持续性和栖息地异质性之间呈现出正相关,而在自然环境中,异性环境中的有性生殖种群更容易被无性生殖所取代[ 77 •• ]。 在所有情况下,无性系最终都胜过有性系,但不清楚这种观察在多大程度上是由于一些无性系的广泛分布或不同无性系划分生态位空间[ 77 •• ]。
挑战和前景
昆虫是了解性别及其丢失原因和影响的重要模型。 随着基因组方法的不断增长的实用性,这些系统将变得更加强大。 但是,存在一些障碍:
首先,昆虫在实验室研究中的易处理程度不同。 有一些发达的系统(例如草蓟马 、粘虫、蚜虫),但对于其它昆虫(如许多寄生蜂),实验室饲养无法维持或未曾尝试过。 评估性别假设可以通过结合实验室和野外研究 来实现 ,后者是推断不同自然条件下 生殖后果的必要条件 [ 78 • ]。
其次,虽然昆虫基因组资源越来越容易获得,例如[ 79 , [80 ],可用无性昆虫数据集的数量仍然相对较少( 表2 )。 此外,据我们所知,只有六份已出版的工作,使用了完整的基因组和/或转录数据集,明确测试有关昆虫性别丢失后果的问题[ 49 •• , 50, 59•• , 61 , 62 , 63• ]。 一个重要的目标应该是测定和生成有性和无性近缘昆虫系统的基因组序列数据。
第三,在某些情况下,可能难以直接将基因组进化模式归因于生殖模式的差异。群体统计学或生态学差异可能会混淆性别丢失的影响。 未来的研究应该优先考虑共存和/或生态相似的有性和无性系近缘昆虫系统。
最后,虽然试图找出有关 无性繁殖的广泛模式有吸引力 , 无性 的原因和后果可能部分取决于转变发生的性别背景[ 81 ]。 因此,应根据具体情况评估性别假设。 例如,无性昆虫中的突变积累可能取决于重组是否持续(无融合生殖与自交孤雌生殖)。另外,在经历不常发生性行为的昆虫(例如,兼性/循环孤雌生殖)中,突变积累可能是特异性的或不存在的,并且这些系统可能不适用于不具有生殖可塑性的其它系统。 增加的基因组资源将能够确定,无性的单独转变具有导致对基因组进化的平行影响相似潜在机制。
https://m.sciencenet.cn/blog-536560-1156211.html
上一篇:Journal Club:昆虫另类遗传系统进化(Benjamin B. Normark,2003)
下一篇:Journal Club:昆虫和螨类孤雌生殖(Normark & Kirkendall 2009)