博文
代谢学人--Nature Metabolism:知否知否,肝EVs控糖高手
||
Nature Metabolism:知否知否,肝EVs控糖高手
撰文 | 郭钰涵 刘梓棋 李姿萱 于柳 李章燕 周文豪 邱瑾
编辑 | 孟美瑶
校对 | 于柳

背景介绍
非酒精性脂肪肝(NAFLD)是肥胖的常见并发症,NAFLD包括一系列肝脏疾病:非酒精性脂肪肝(NAFL,肝脏脂质积聚>5%)和非酒精性脂肪性肝炎(NASH)。NAFLD与胰岛素抵抗、血脂异常、2型糖尿病(T2D)和冠心病密切相关,而发展为NASH则会增加肝纤维化水平、肝硬化水平、肝脏失代偿(小编注:肝脏失代偿是指慢性肝病在病情发展到一定程度时,肝脏功能逐渐受损,无法维持正常的生理功能,导致出现明显的代偿障碍和症状。肝脏失代偿通常是肝硬化晚期的表现,也是严重的肝功能不全的表现之一。肝脏失代偿的表现包括但不限于:1)腹水(腹腔内积液):由于肝功能受损,无法正常合成蛋白质,导致血浆胶体渗透压降低,血管内液体渗出增多,最终在腹腔内积聚形成腹水。2)肝性脑病:肝功能不全导致肝脏无法有效清除体内毒素,使得毒素如氨等物质在体内蓄积,影响大脑功能,出现认知障碍、意识模糊等症状。3)肝昏迷:是肝性脑病的严重表现,患者表现为昏迷状态,需要紧急治疗。4)出血倾向:肝脏失代偿时,凝血功能受损,易出现出血倾向,如鼻出血、消化道出血等。5)肝肾综合征:肝脏失代偿时,肾脏功能也可能受到影响,出现肝肾综合征,表现为肾功能减退、尿量减少等症状。)等与肝脏相关的疾病发生率。据统计,世界上约25%的人口患有NAFLD,这其中有20%的病例被归类为NASH。值得注意的是,肥胖患者的NAFLD发病率约为75%,T2D患者的发病率为56%(小编注:肥胖患者的NAFLD发病率75%来自于一项澳大利亚的队列研究,该研究纳入了216名BMI = 45.9±8.9kg/m2的患者,NAFLD总体患病率为74.1%(Ooi, G. J. et al. . Obes. Surg. 29, 99–108 (2019).)。而T2DM中56%的NAFLD患病率来自2019年的一项荟萃分析,该研究纳入80项有关T2DM患者NAFLD患病率的研究,统计分析发现T2DM患者的全球NAFLD患病率估计为55.48%(95%CI 47.26-63.67%)(Younossi, Z. M. et al. . J. Hepatol. 71, 793–801 (2019))。
目前,NAFLD与糖尿病密切关联的原因尚不明确。研究表明,NAFLD与肥胖患者外周组织中的胰岛素抵抗有关,这暗示肝脏分泌的体液因子会影响远处组织对胰岛素的敏感性。NAFLD患者肝源性蛋白质(肝因子)可通过“经典分泌”途径从肝脏释放出来:即含有N-末端信号肽的肝因子在内质网内发生糖基化后,被蛋白复合物II包被的囊泡运输到高尔基体,经过进一步的加工到达细胞膜,将蛋白质释放到细胞外,通过自分泌/旁分泌或内分泌到达靶细胞,并发出信号。虽然经典的蛋白质分泌途径是组织通讯的标志,但肝脏分泌的80-90%的蛋白质不含信号肽,这表明通过其他机制分泌的蛋白质可能是组织间通讯和代谢控制的媒介。
细胞外囊泡(EVs)是一种纳米尺寸的囊泡,含有脂质、蛋白质和核酸,可转运到受体细胞并引发功能反应。目前关于肝源性EVs调节其他细胞和组织生理过程的研究相对较少,关于NAFLD进展如何影响肝源性EVs组成的变化目前也尚未明确。研究表明,从早期肥胖小鼠体内分离出来的肝源性EVs可通过其包含的miR-3075增强受体小鼠的胰岛素敏感性。相反,从慢性肥胖小鼠中分离出的肝源性EVs引起胰岛素抵抗,这或与外周组织的促炎性巨噬细胞激活(小编注:研究发现长期(16周)HFD饮食小鼠的肝源性EVs处理瘦小鼠后,会引起受体瘦小鼠的脂肪组织、肌肉组织和肝脏促炎反应通路激活,进一步通过流式分析发现受体瘦小鼠eWAT中的促炎巨噬细胞被激活。Ji, Y. et al. Nat. Metab. 3, 1163–1174 (2021).)有关。虽然上述研究为阐明肝源性EVs如何通过促进组织适应性以影响血糖控制奠定了重要基础,但非肝实质细胞(例如Kupffer细胞、肝星状细胞)的EVs是否会影响这些过程,而这些过程在从NAFL到早期NASH的过程中是否会发生改变,以及肝源性EVs是否会激活急性细胞信号传导事件并快速调节靶组织中的代谢反应等问题目前尚不清楚。此外,最近的研究表明,microRNA(miRNA)不是EV的主要成分,EV的其他成分,如蛋白质,可能是EV介导细胞通讯的重要调节因子。鉴于EV携带多种胞内蛋白和跨膜蛋白,并且EV的产生会受到激素和营养物质的影响,因此研究人员假设肝源性EVs可随营养物质利用率的增加而分泌增多,并作用于受体细胞以调节新陈代谢。
本研究中,研究人员使用蛋白质组学来鉴定小鼠从健康肝脏向NAFL和NASH转变过程中肝源性EVs的蛋白质组成变化。研究表明无论肝脏处于何种病理状态,来源于小鼠/人肝脏的EVs都能通过增加胰岛素分泌和增强葡萄糖有效性(GE)(小编注:GE是指葡萄糖通过不依赖胰岛素的机制影响自身代谢的能力,包括促进葡萄糖利用和抑制葡萄糖合成两个途径。),来快速改善血糖控制,并且该作用与胰岛素敏感性的变化无关。此外,上述反应对肝源性EVs是特异性的,并且依赖于EVs跨膜蛋白的存在。在健康状态下,充足的葡萄糖供应会增强肝源性EVs的分泌,而在胰岛素抵抗状态下分泌则受到抑制,这表明肝源性EVs能响应波动的血糖水平从而协助机体快速调节血糖。

敲黑板啦!
1、肝源性EVs可增加葡萄糖耐受能力,并促进胰岛素分泌
2、肝脏感应葡萄糖促进肝源性EVs的分泌
3、肝脏分泌的EVs不调节胰岛素敏感性
4、肝源性EVs增强骨骼肌葡萄糖有效性(GE)并调节其细胞信号传导
5、肝源性EVs促进胰岛中葡萄糖刺激的胰岛素分泌
6、人肝源性EVs可改善血糖控制
研究结果
1. 细胞外囊泡分泌与NAFLD进展
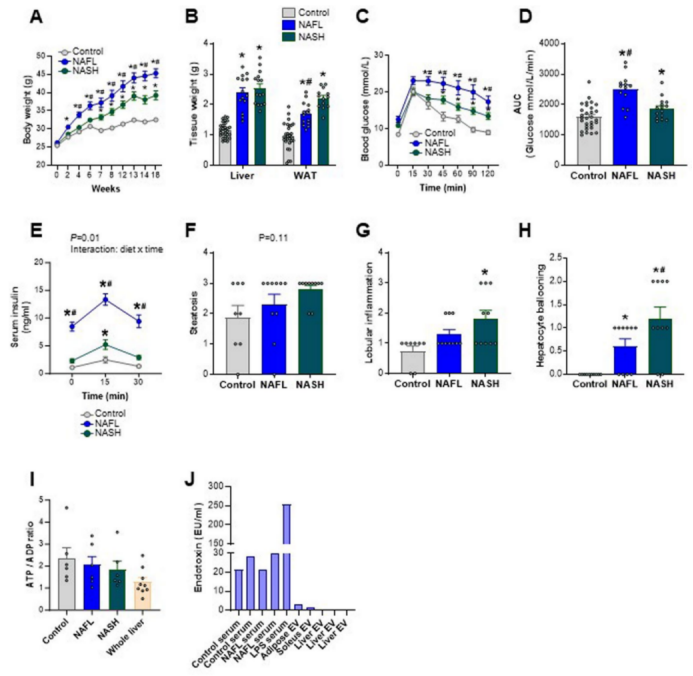
NAFLD和胰岛素抵抗是常见的合并症;已有多项研究表明,肝脏分泌因子会影响胰岛素的作用和血糖控制。因此,研究人员假设与对照组相比,NAFL和NASH中肝源性EVs的组成发生了改变,并且这些EVs削弱了小鼠的血糖控制能力。研究人员首先在饮食诱导的NAFL和NASH小鼠模型(小编注:对照组饮食:12%脂质、25. 8%的蛋白质和 62.2% 的碳水化合物,NAFL饮食:高脂饮食,43%脂质、21%的蛋白质和 37% 的碳水化合物)。NASH饮食:高脂/高果糖饮食,2% 胆固醇饮食,39.8%脂质、20%的蛋白质和 40.2% 的碳水化合物,共喂养20周。)中进行验证,结果发现与对照组相比,喂食NASH和NAFL饮食小鼠的体重、肝脏质量和附睾脂肪组织(eWAT)质量增加(扩展数据图1a、b)。与正常喂养小鼠相比,喂食NAFL饮食和NASH饮食的小鼠均表现出严重的葡萄糖不耐受(扩展数据图1c,d)和高胰岛素血症(扩展数据图1e)(小编注:这里使用的是多因素方差分析,分析了不同饮食,不同时间点以及饮食与时间点的交互项对血清胰岛素的影响。diet x time代表饮食与时间点的交互项.)。
肝脏组织病理学结果显示,与对照小鼠相比,NAFL和NASH小鼠的肝脏发生明显的脂肪变性(图1a),甘油三酯水平增加(图1b)。与对照组相比,NAFLD活性评分(NAS)(小编注:NAS评分是通过对肝脏组织镜检分析,根据肝脏炎症(气球样变和小叶内炎症)、坏死和纤维化等方面的严重程度进行评分,其得分范围为0-8分,得分越高则NAFLD越严重。)在NAFL小鼠中显著增加,在NASH小鼠中进一步增加(图1c,扩展数据图1f–h)。与对照小鼠相比,NAFL和NASH小鼠中的血清ALT(肝损伤的标志物)水平更高(图1d)(小编注:通过比色酶法检测血清中ALT的浓度;该方法来源于国际临床化学联合会,基本原理为ALT催化底物丙氨酸产生丙酮酸;而丙酮酸在NADH存在的条件下,可被乳酸脱氢酶的催化下产生乳酸;其中实验反应体系中需要添加丙氨酸和NADH等;通过光谱仪检测数值变化,最后酶活性主要通过NADH的消耗速度来间接反映。)纤维化是NAFLD发生发展过程中的一个特征,尽管NASH小鼠的肝脏中有几个促纤维化基因表达上调(例如,Tgf1β、Timp1、Cd11;图1e),但在NAFL和NASH小鼠中均没有观察到组织学上的纤维化表型(图1a)。总之,以上模型概括了NAFL和早期NASH的特征,且未发现肝脏出现明显的纤维化(小编注:推测本文应该是想强调该NASH模型为早期NASH模型,因为从促纤维化基因表达以及NAFLD活性评分(NAS)的角度看,该NASH模型较NAFL相比会更严重些。)。
为了确定NAFLD对肝源性EV分泌的影响,研究人员从小鼠身上切除肝脏,制备300μm的精密切片,并在含氧及富含营养的缓冲液中孵育(小编注:本实验是将整个小鼠肝叶或人活检组织切片300um厚度后进行培养。也有研究利用脂肪组织,使用插入式细胞培养皿模拟体内环境培养350um厚度的人脂肪组织切片,并通过免疫荧光检测 Perilipin A评估脂肪细胞的活力。Schopow N, et al. PLoS One. 2020)。考虑到肝损伤和应激的可能性,研究人员检测了供体小鼠的肝切片和速冻全肝中的ATP:ADP比率(小编注:细胞内的ATP:ADP比率是细胞活力的重要指标之一。ATP是细胞内的能量储备分子,而ADP是ATP水解后形成的产物。细胞活力的变化通常会导致ATP:ADP比率的变化,因为细胞需要ATP来进行各种生物学过程。因此通常通过监测ATP:ADP比率可用来评估细胞的代谢状态和活力水平),发现该比率在各组之间没有差异,表明肝切片是具有活力的(扩展数据图1i)。研究人员将肝切片孵育16小时后收集肝脏分泌因子,并使用差速离心技术分离和纯化EVs。纳米颗粒跟踪分析(NTA)显示,大多数EVs的尺寸都符合预期(约130 nm;图1f);低温透射电子显微镜(cryoTEM)显示,EV由双层脂质和球形结构组成(小编注:冷冻透射电镜可降低样本损伤,能够更加准确的反映EV结构)。(图1g)免疫印迹分析实验证实了TSG101、CD63、CD9、HSP70和CD81等EV蛋白的存在(图1h)。而EV中不表达calnexin蛋白进一步证实了EVs的纯度(calnexin是一种特异性定位于内质网的蛋白质)(图1h)。此外,肝源性EVs几乎没有内毒素污染,其含量比在小鼠血清中检测到的内毒素含量低约50倍(扩展数据图1j)(小编注:文中分离的是NAFLD小鼠的肝组织EV,有研究表明,肠道菌群失调驱动的肠道屏障功能障碍是NASH发展的重要促成因素(Nat Microbiol . 2023 Aug;8(8):1534-1548. )(Life Sci . 2023 Jun 1:322:121685. )。内毒素是革兰氏阴性菌的细胞壁组分,也称为LPS,肠道菌群可利用其代谢物诱发糖脂毒性、氧化应激和肠道屏障损伤,肠道屏障破坏导致过度生长的细菌及其产物易位到粘膜和循环中,内毒素经门静脉进入肝脏,从而通过激活肝脏中的炎性细胞诱导炎症反应,已有研究表明,NAFLD个体的全身LPS浓度显著升高(J Inflamm (Lond) . 2010 Mar 30:7:15. )。推测从体外肝组织切片中提取EV可避免肠道中内毒素对EV质量的干扰)。
接下来,作者探究了NAFL和NASH对肝源性EVs的影响。NTA分析表明,与对照组和NAFL小鼠相比,NASH小鼠肝脏分泌的EVs浓度增加(图1i)。研究人员发现,以每毫克肝脏切片为单位时,NAFL和NASH小鼠分泌的EVs中的蛋白质丰度比对照小鼠低(图1j);然而,当以肝脏总质量为量化单位时,各组之间分泌的EVs中的蛋白质量相似(图1k)。总之,这些数据表明,NAFL和NASH中的EVs分泌增加,但分泌的EVs中的总蛋白质水平与对照组相似。
扩展数据图1. 饮食诱导的葡萄糖不耐受情况和肝源性EV纯度测试
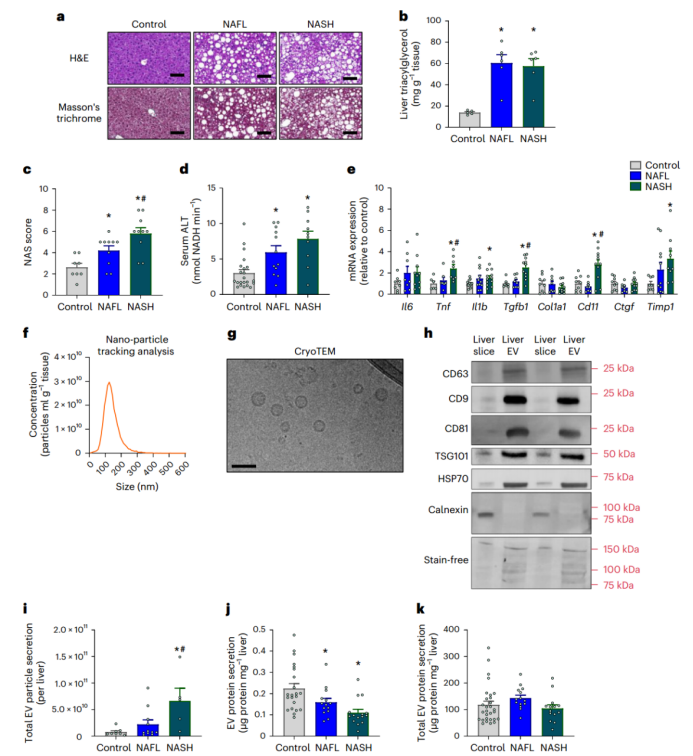
图1. NAFL和NASH小鼠表型以及小鼠肝脏分泌的EV分离物验证
2. NAFL和NASH小鼠肝源性EV蛋白质组发生改变
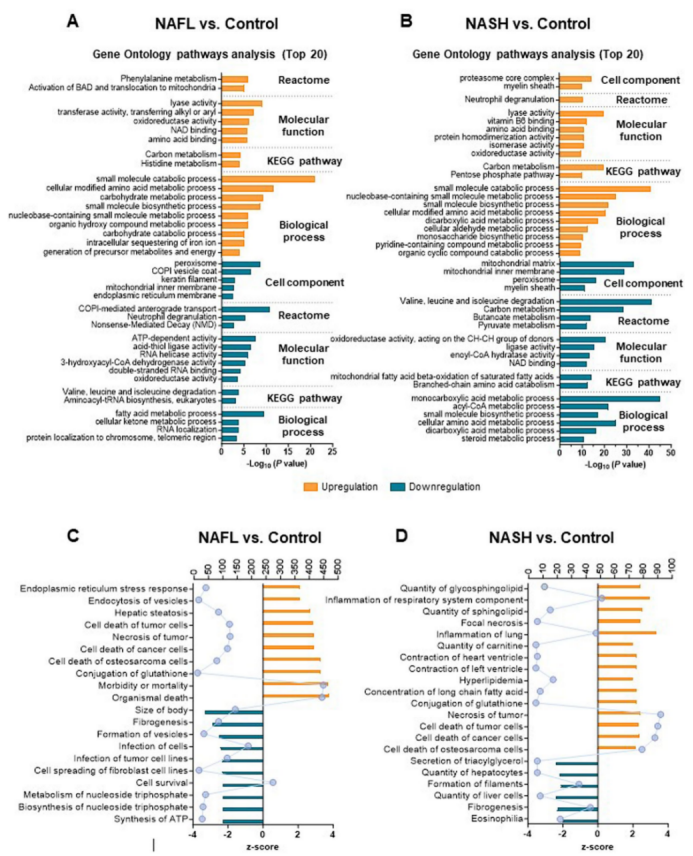
为了研究来源于肝脏组织的EVs组成在NAFL和NASH状态下是如何发生变化的。因此,研究人员通过液相色谱-串联光谱法(LC–MS/MS)蛋白质组学,鉴定了肝源性EVs中包含的1779种蛋白质;其中1688种蛋白质在对照组、NAFL和NASH组中都表达(图2a)。主成分分析(PCA)(图2b)进一步分析发现,与对照组相比,NAFL小鼠的肝源性EVs中,有91种蛋白水平出现上调,66种蛋白水平出现下调(图2c);而与对照小鼠相比,患有NASH小鼠的肝源性EVs中则有222种蛋白水平出现上调,249种蛋白水平出现下调(图2d)。值得注意的是,NAFL和NASH小鼠的蛋白质组之间没有差异(小编注:如前文模型评估数据显示,NAFL和NASH小鼠在组织学方面无显著差异,但从促纤维化基因、NAFLD活动性评分和血清ALT水平方面来看,NASH小鼠要比NAFL小鼠更严重一些,也表明两者之间存在差异,但并未到达组织学可观测的水平。推测,这或许是作者强调本文的NASH模型处在早期阶段的原因。因此,后续作者认为肝源性EV蛋白组成的变化是NAFLD进展的早期表现)(图2e),表明肝源性EV蛋白组成的变化是NAFLD进展的早期表现(小编注:这里的蛋白质组数据所反映的NAFL和NASH组间无差异,但均与对照组有差异,突出这里的蛋白组成差异主要是从NAFL阶段开始的,即使继续进展到早期NASH但并未显著改变EV蛋白组成;由此得出的“肝源性EV蛋白组成的变化是NAFLD进展的早期表现”的结论)。通路富集分析表明,NAFL和NASH小鼠肝EVs中与碳水化合物代谢和小分子分解代谢过程相关的蛋白水平上调,与线粒体β-氧化和ATP依赖的脂肪酸代谢相关的蛋白水平下调(扩展数据图2a,b)。进一步的分析表明,NAFL和NASH小鼠肝EVs含有表征糖酵解和糖异生、糖原代谢、丙酮酸代谢和三羧酸(TCA)循环水平上调有关的蛋白质,以及表征脂质运输、延伸、合成和氧化过程水平下调有关的蛋白质(图2f,g)。研究人员使用IPA(Ingenuity Pathway Analysis)分析法进一步分析发现,来自NAFL或NASH小鼠肝源性EVs中的蛋白质与严重的肝脂肪变性、高脂血症和癌细胞死亡、肝细胞数量减少、ATP合成及纤维化有关(扩展数据图2c,d)。总之,这些数据强调了NAFLD中肝源性EVs蛋白质组的改变,并且这些蛋白质与碳水化合物的各种代谢途径的上调有密切联系。

图2. NAFL和NASH小鼠的肝源性EV蛋白质组
扩展数据图2. 肝脏细胞外囊泡蛋白质组对NAFL或NASH反应的通路分析
3. 肝源性EVs增加葡萄糖耐受能力,并提高胰岛素分泌水平
由于NAFL和NASH会引起肝源性EVs蛋白质组的显著变化,且与葡萄糖代谢紧密相关。因此,研究人员进一步探究了肝源性EVs对机体调控血糖能力的影响。首先,研究人员给正常小鼠腹腔或者静脉注射来自NAFL小鼠或NASH小鼠的EVs ,或者给患有NAFL或NASH的小鼠注射源来自健康小鼠的EVs,1h之后对进行代谢相关检测。令人惊讶的是,无论EVs供体是哪一种小鼠,其肝脏分泌的EVs都大大改善了所有小鼠的全身血糖耐受能力。即,在口服葡萄糖后,注射肝源性EVs的对照组、NAFL和NASH雄性小鼠的血糖水平均降低,曲线下葡萄糖面积(AUC)减少。(图3a–c和扩展数据图3a–c),而在接受腹腔注射肝源性EVs的对照雌性小鼠中也观察到类似的结果(扩展数据图3d)。此外,研究人员还探究了静脉注射肝源性EVs的效果,发现静脉注射来自NAFL小鼠肝源性EV之后,正常小鼠的血糖控制能力也得到了显著改善(扩展数据图3e),并且腹腔和静脉注射肝源性EVs后均能显著增加小鼠血清胰岛素的水平(图3d–f和扩展数据图3f)。为了确定以上作用是肝源性EVs产生的独有效应还是所有EVs的共同特征,研究人员分别给小鼠注射了空脂质体、脂肪分泌的EVs或血清衍生的EVs,并进行口服葡萄糖耐量测试(GTT),结果发现各组的葡萄糖耐量(扩展数据图3g–i)和血清胰岛素水平(扩展数据图3j)没有差异,表明肝源性的EVs在血糖控制方面具有特异性。

图3. 肝脏分泌的EVs改善血糖控制

扩展数据图3. 小鼠对各种来源的细胞外囊泡和葡萄糖刺激的细胞外囊泡分泌的反应的葡萄糖耐受性测试
4. 肝脏通过感应葡萄糖增加肝源性EVs的分泌
基于肝源性EVs可改善葡萄糖刺激状态但非基础状态下的血糖控制(小编注:基础状态下的血糖控制指的是注射生理盐水组(图3a-c),血糖控制没有得到改善,而给正常小鼠腹腔或者静脉注射来自NAFL小鼠或NASH小鼠的EVs ,或者给患有NAFL或NASH的小鼠注射源来自健康小鼠的EVs之后,1h之后给小鼠口服葡萄糖进行代谢相关检测,发现口服葡萄糖后,各组小鼠的血糖水平降低,血糖控制能力得到改善,因此注射EV改善的是“葡萄糖刺激状态下的血糖控制”) 。研究人员提出高血糖能刺激肝脏分泌EVs的假设。接下来,研究人员用胰岛素(10 nM)、胰高血糖素(20 pg ml−1)或去甲肾上腺素(15 pg ml−1)处理来源于健康小鼠的肝组织,并施加低(2 mM)或高(20 mM)葡萄糖刺激;之后从培养基中收集EVs。在高糖(HG)刺激下,肝脏分泌的EVs浓度和总EVs蛋白含量增加(图3g,h);且胰岛素、胰高血糖素和去甲肾上腺素均不会影响肝源性EVs的分泌(扩展数据图3k,l)。蛋白质组学分析显示,肝源性EVs的蛋白质组在LG和HG条件下没有差异(扩展数据图3m),这表明在蛋白质组成没有变化的情况下,HG刺激会促进肝源性EVs的释放。为了探究体内实验是否也能得到类似的结果,研究人员评估了禁食状态和口服葡萄糖后小鼠血清中肝源性EVs的量(小编注:作者使用成像流式细胞术分析来鉴定典型EVs标记物CD9、CD63的存在,并且通过与肝脏特异性蛋白唾液酸糖蛋白受体1(ASGPR1,编码基因为Asgr1共染来鉴定肝源性EVs。ASGPR1是存在于肝细胞的窦状隙面的一类跨膜糖蛋白,被认为是肝脏表面的标志物(Development . 2016 May 1;143(9):1475-81.),上文提到的CD9和CD63是所有EVs的maker,作者用ASGPR1和CD9、CD63共染来确定EVs来自肝脏,所以,可认为ASGPR1是肝源性EVs上的maker。此外,GPRC5A、AGER被认为是肺特异性EVs标志物(J Extracell Vesicles . 2022 May;11(5):e12223. )。结果发现,与禁食小鼠相比,葡萄糖给药后健康小鼠血清中肝源性EVs的数量增加(图3i、j和扩展数据图3n-q)。然而,NAFL小鼠对葡萄糖刺激的肝源性EVs分泌具有抗性(图3k和扩展数据图4a–d)。以上数据表明,肝脏能感知葡萄糖供应的增加并促进EVs的分泌,而对葡萄糖刺激EVs分泌的抵抗是NAFL的一个特征。

扩展数据图4. 小鼠及分离肝组织对各种处理产生的细胞外囊泡的糖耐量测试
5. 改善血糖控制需要完整的肝源性EV
EVs中含有蛋白质、脂质和核酸,这些物质被输送到受体细胞并影响细胞功能。为了确定单独的EVs成分是否足以改善血糖控制,研究人员使用超声处理破坏EVs结构后,将其注射到小鼠体内,结果显示,在对照或NAFL小鼠中,注射经超声处理过的EVs的小鼠血糖(图3l和扩展数据图4e)和胰岛素水平(图3m)与注射生理盐水的小鼠没有差异,这表明肝源性EVs必须保持完整,其生物活性成分才能诱导上述的血糖效应(图3a–c)。此外,细胞受体蛋白为重要媒介,可通过受体-配体结合、融合或内吞作用实现EVs-靶细胞相互作用。接下来,研究人员发现,使用蛋白酶K消化除去EVs表面蛋白会削弱EVs改善血糖的效果(图3n、p和扩展数据图4f、g),并抑制血清胰岛素水平的升高(图3o、q);而除去EVs表面蛋白不会改变EVs的形态(扩展数据图4h)或大小(扩展数据表4i)。因此,EVs表面蛋白是肝源性EVs增加胰岛素分泌和改善小鼠血糖控制所必需的。
研究表明,大脑通过抑制肝脏葡萄糖输出、促进胰岛素分泌和骨骼肌葡萄糖摄取来调节血糖水平。脂肪来源干细胞的EVs可以与大脑中的小胶质细胞进行通信(小编注:有研究发现脂肪源EVs中的miRNA可以靶向小胶质细胞中的PTEN和STAT1通过调节小胶质细胞极化来减少脑缺血带来的损伤。参考文献:Hu X, Pan J, Li Y, et al. Stem Cell Res Ther. 2022),这暗示肝源性EVs有可能通过直接作用于大脑来调控血糖。于是研究人员在GTT前1-3小时通过侧脑室(ICV)注射,将生理盐水或NAFL小鼠的肝源性EVs(16 μg)直接递送至中央脑室系统,结果发现,与注射生理盐水相比,将肝源性EVs直接注入大脑不会改变血糖耐受能力(扩展数据图4j)和血清胰岛素水平(扩展数据表4k),表明肝源性EVs是通过与外周组织直接接触从而影响血糖耐受能力和胰岛素分泌水平。
6. 肝脏分泌的EVs不调节胰岛素敏感性
由于肝源性EVs的长期给药可改善外周胰岛素功能(小编注:该结论来自于参考文献引用,该研究发现肝细胞会分泌富含miR-3075的外泌体,从而增强外周胰岛素敏感性。参考文献:Ji Y, Luo Z, Gao H, et al. Nat Metab. 2021),研究人员推测,肝源性EVs可能通过增强胰岛素敏感性来降低血糖。出乎意料的是,无论在对照还是NAFL小鼠中,肝脏分泌的EVs均不会改变ITT水平(图4a,b)。此外,在生理盐水条件或胰岛素刺激条件下,肝源性EVs处理均不会影响原代肝细胞的葡萄糖输出水平(图4c),也不会影响对照或NAFL小鼠的离体骨骼肌,白色和棕色脂肪组织的葡萄糖摄取能力(图4d–f和扩展数据图4l–n)。高胰岛素-正常血糖钳夹实验对对照小鼠的胰岛素敏感性评估结果也支持上述结论(小编注:高胰岛素-正常血糖钳夹实验是至今为止公认的测定机体胰岛素抵抗的金标准。通过给予外源性的胰岛素,使机体处于高胰岛素状态,且这一胰岛素水平能抑制内源性葡萄糖的产生,同时输注葡萄糖,使血糖浓度被控制在基础水平,达到外源性胰岛素与葡萄糖代谢的平衡。在这一稳定状态下,葡萄糖输注率等于葡萄糖在体内被胰岛素敏感组织(脂肪、肌肉)的吸收代谢率,因此,可以根据葡萄糖输注率来反映外周组织对外源性胰岛素的敏感性。);结果显示接受生理盐水或NAFL EVs的小鼠在禁食状态下和正常血糖钳夹实验下血糖浓度无显著差异(扩展数据图4o);并且在钳夹过程中,所有小鼠的血浆胰岛素水平都升高(扩展数据图4p)。钳夹期间的葡萄糖输注速率(图4g)反映了全身胰岛素敏感性,两组之间没有差异。总之,这些独立的实验表明,肝脏分泌的EVs并不能通过增强胰岛素敏感性来快速改善血糖控制。
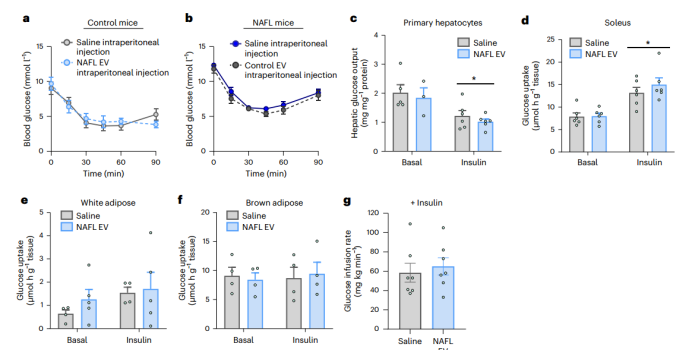
图4. 肝脏分泌的EVs不调节胰岛素敏感性
7. 肝源性EVs增强骨骼肌的GE
GE是指葡萄糖通过不依赖胰岛素的机制影响自身代谢的能力,可占人类葡萄糖利用率的45-65%。为了探究EVs是否能调节GE,研究人员在胰岛素缺乏的短期模型中来检测肝源性EVs的血糖控制能力。在小鼠GTT前2周,用低剂量链脲佐菌素(STZ)(40 mg kg−1)处理小鼠以降低胰岛素的水平。与健康对照小鼠相比,STZ处理的小鼠出现严重的葡萄糖不耐受,但给STZ处理小鼠注射来自健康小鼠的肝源性EVs一个小时之后进行GTT实验发现其血糖控制能力得到显著改善(图5b和扩展数据图5a)。另一方面,向具有胰岛素抵抗的对照小鼠和NAFL小鼠腹腔注射葡萄糖后,再分别使用来自NAFL小鼠的肝源性EVs处理对照小鼠,或者使用来自对照小鼠的肝源性EVs处理NAFL小鼠,都能明显改善对照小鼠和NAFL小鼠的血糖控制能力(图5c,d)。总之,这些数据表明,肝源性EVs可通过增加GE来改善血糖控制(小编注:本段的第一个实验使用了STZ小鼠,排除了EVs通过促进机体分泌胰岛素改善血糖控制能力的可能性,第二个实验使用了胰岛素抵抗小鼠,排除EVs通过快速改善胰岛素敏感性来改善血糖控制能力的可能性,从而说明EVs可通过增加GE来改善血糖控制)。
为进一步佐证上述结论,研究人员给健康小鼠注射NAFL小鼠的肝源性EVs,结果发现,与对照组相比,肝源性EVs给药后2h内,小鼠全身碳水化合物氧化水平增加了两倍以上 (图5e),且与脂肪氧化水平(扩展数据图5b)、能量消耗增加(扩展数据图5c)以及体力活动(扩展数据图5d)无关。接下来,研究人员在体外进一步探究NAFL来源的EVs与骨骼肌中GE(小编注:上文有实验显示在正常的葡萄糖浓度(Method中为10uM)下,与对照组相比,在胰岛素处理的情况下,EVs处理未影响骨骼肌的葡萄糖摄取,主要是为了说明胰岛素敏感性未发生改变。这里的实验是在高糖情况下做的,因为葡萄糖有效性的测量需要在高浓度葡萄糖刺激条件下进行(图5f显示在20uM情况下才有差异)之间的关系),结果发现,在高浓度的2-脱氧葡萄糖(2-DG)条件下,NAFL小鼠肝源性EVs(10 μg ml−1)能促进完全分化的C2C12细胞的葡萄糖摄取(图5f),并且在分离的小鼠比目鱼肌中也得到了同样的结果(图5g)。以上数据表明,肝源性EVs能够直接促进骨骼肌中的GE,与胰岛素或其他内分泌因素无关。
为了确定肝源性EVs促进GE的机制,研究人员在STZ小鼠中进行研究,并重点研究了骨骼肌中的GLUT4易位。研究人员首先给小鼠注射生理盐水或者正常小鼠的肝源性EVs(30 µg),并在1 h之后用水或葡萄糖灌胃,结果发现葡萄糖并不会刺激胰岛素的分泌,这表明STZ造模成功。(扩展数据图5e)。与用生理盐水+溶剂和生理盐水+葡萄糖处理的小鼠相比,用肝源性EVs+葡萄糖处理的小鼠质膜上的GLUT4蛋白水平更高(图5h),微粒体上的GLUT4蛋白相应减少(小编注:微粒体是当细胞被匀浆破碎时, 内膜系统的膜结构破裂后自己重新封闭起来的小囊泡。由于GLUT4蛋白可在细胞膜系统间易位;因此在GLUT4蛋白水平更高的条件下,内膜系统的GLUT4蛋白水平反而降低,间接说明GLUT4蛋白易位到了细胞膜上,发挥葡萄糖摄取功能)(图5i)。各组的质膜caveolin 3蛋白(介导细胞内吞作用)含量没有差异(图5h和扩展数据图5f)。以上结果表明肝源性EVs可通过增加质膜GLUT4易位,增强骨骼肌中的GE。

图5. 肝源性EVs通过增强GE改善血糖控制

扩展数据图5. 肝细胞外囊泡在调节葡萄糖有效性中的作用
拓展阅读
葡萄糖有效性(GE)
葡萄糖有效性是指在葡萄糖浓度快速上升时,葡萄糖可通过不依赖胰岛素的机制影响自身代谢的能力,主要包括促进葡萄糖利用和抑制葡萄糖合成两个途径,从而达到降低外周葡萄糖浓度的目的。目前已经在动物实验上证明单独使用葡萄糖并在基础胰岛素水平持续较低的情况下,可促进机体肝脏的葡萄糖摄取能力和糖代谢水平,并且可以刺激肝内糖原合成,同时抑制肝糖原分解。另外有体外实验表明,高葡萄糖刺激可以在体外以非胰岛素依赖的形式,促进C2C12肌管内的Glut4易位到细胞膜上并促进糖摄取。
参考文献:[1] Petersen KF, Laurent D, Rothman DL, Cline GW, Shulman GI. J Clin Invest. 1998[2] Galante P, Mosthaf L, Kellerer M, et al. Diabetes. 1995[3] Kruszynska YT, Home PD, Alberti KG. Diabetes. 1986
8. 肝源性EVs调节骨骼肌细胞信号传导
为了深入了解肝源性EVs调节GE的机制,研究人员对分离的比目鱼肌分别给予低糖(LG)或高糖(HG)刺激;以及在HG刺激下,使用来自NAFL小鼠肝源性EVs(10 μg ml−1)进行处理,并对比目鱼肌进行磷酸化蛋白质组学分析。研究人员总共分析了2247种蛋白质中的13140个磷酸位点。与LG相比,HG刺激导致27个磷酸化位点上调,23个磷酸化位点下调(图6a)。与单独的HG刺激相比,在HG刺激的情况下结合NAFL小鼠的肝源性EVs处理比目鱼肌,会导致61个磷酸化位点的上调和48个磷酸化位点的下调(图6b)。其中包括蛋白质STIM1(S524)的磷酸化水平增加(STIM1是钙稳态的调节因子,在小鼠进食的情况下,小鼠肝脏中STIM1磷酸化水平会升高),这一结果支持了STIM1在调节GE中可能发挥的作用。肝源性EVs改变的磷酸化位点与单纯HG调控的磷酸化位点完全不同,这表明葡萄糖依赖性和葡萄糖非依赖性途径可能协同介导EVs对GE的影响。生物学过程的通路分析表明,不同条件处理的比目鱼肌的共同信号传导通路差异较大,而在HG刺激下,对照组和肝源性EVs处理组之间的信号传导通路差异最大(扩展数据图5g,h)。然而,有一些蛋白质受到HG和EVs的共同调节,但值得注意的是,调节发生在不同的磷酸化位点(图6c,d)。这包括蛋白PROB1(S396、S407、S398、S451)和TNS1(S1427、S963)中磷酸化位点的下调,以及NACA(S1914、S1491)、CAMK2b(S397、T400)和PDHA1(S295、S300)中磷酸位点的上调。而CAMK2b和PDHA1分别是已知的葡萄糖摄取和氧化的调节因子。
生物信息学分析表明,HG刺激会激活骨骼肌的钙敏感通路(扩展数据图5g),肝源性EVs进一步激活了钙相关的各种信号事件,包括丝氨酸和苏氨酸特异性蛋白激酶Ca2+/钙调素依赖性蛋白激酶II(CaMKII)。因此,研究人员用丹曲洛林抑制肌浆网的钙释放,并用细胞渗透性抑制剂KN-62抑制CaMKII活性,然后评估EVs介导的GE。结果发现,在HG刺激下,EVs没有进一步提高丹曲洛林(抑制肌浆网钙的释放)和KN-62(钙调蛋白依赖性蛋白激酶 II (CaMK-II) 抑制剂)处理过的离体小鼠骨骼肌的葡萄糖摄取水平(图5f,g)(图6e,f)。值得注意的是,在无EVs的LG或HG条件下,丹曲洛林和KN-62对葡萄糖摄取均没有影响(扩展数据图5i)。此外,在C2C12肌管中,丹曲洛林和KN-62也消除了EVs介导的葡萄糖摄取增加(图6g,h)。总之,这些数据证明在HG和肝源性EVs干预下,骨骼肌会做出不同的信号传导反应,其中肌浆网钙释放和CaMKII激活在EVs介导的GE中扮演着重要角色。

图6. 肝源性EVs激活与钙代谢相关的细胞信号传导
拓展阅读
Ca2+和骨骼肌GLUT4转运
在骨骼肌中,Ca2+能激活Ca2+/钙调蛋白复合物与CaM 结合域,导致Thr286磷酸化,从而激活 CaMKII。CaMKII 激活能活化HDAC,增加MEF2基因转录水平;之后MEF2与HAT和其他转录因子共同促进GLUT4的表达,并促进GLUT4转运,影响葡萄糖摄取能力。
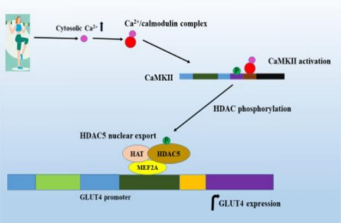
参考文献:[1] Joseph, Jitcy S et al. EXCLI journal vol. 20 386-399. 17 Feb. 2021
9. 肝源性EVs直接刺激胰葡萄糖刺激的胰岛素分泌
考虑到肝源性EVs对小鼠胰岛素分泌的影响,研究人员接下来研究EVs是否可以直接作用于胰腺来增加葡萄糖刺激的胰岛素分泌(GSIS)。与前面的体内研究结果(肝源性EVs增加GSIS(图3d–f))一致,用HG处理分离的小鼠胰岛,随后用NAFL肝源性EVs处理后发现,小鼠胰岛素分泌水平增加两倍(图7a)。然而,当胰岛暴露于用蛋白酶K处理的EVs时,这种促进效应消失,暗示EVs跨膜蛋白可以调节肝源性EV-胰岛相互作用以及胰岛素分泌(图7a)。
为了进一步探究肝源性EVs促进GSIS的可能机制,研究人员对胰岛素的分泌途径开展了系统性研究(图7b)。已知葡萄糖氧化和随后ATP:ADP比率的升高是K+通道抑制和胰岛素分泌的关键事件。在HG刺激的情况下,肝源性EVs能促进葡萄糖氧化水平(图7c),但并没有增加ATP:ADP比率(图7d)。此外,研究人员使用甲苯丁酰胺(一种增加胰岛素分泌的K+通道抑制剂)或二氮氧化物(一种抑制胰岛素分泌的K+通道激活剂)处理胰岛,随后再用肝源性EVs处理所有组,结果发现,与对照组相比,药物处理组的胰岛素分泌水平没有差别(图7e,f和扩展数据图6a,b),表明肝源性EVs通过ATP非依赖性事件调控胰岛素分泌(小编注:ATP依赖性的胰岛素分泌机制,依赖于ATP水平的升高作为信号,本研究结果发现肝源性EVs并没有增加ATP:ADP比率,暗示肝源性EVs通过ATP非依赖性事件调控胰岛素分泌。此外,K+通道为ATP依赖性的胰岛素分泌途径中的重要节点,同样不影响肝EV对胰岛素分泌。综上得出EVs通过ATP非依赖性事件调控胰岛素分泌的结论);这包括精氨酸增加质膜的去极化(图7g和扩展数据图6c)和丙氨酸激活K+通道下游的电压敏感钙变化(图7h和扩展数据图6d)。此外,研究人员用exendin 4(GLP1受体的有效肽激动剂;能通过环AMP激活GLP1依赖的胰岛素分泌)处理暴露于肝源性EVs的胰岛,肝源性EVs增加了GLP1依赖性胰岛素分泌(图7i和扩展数据图6e),这进一步支持了HG刺激下肝源性EVs介导ATP非依赖性效应的可能性。在上述实验中,细胞胰岛素含量在不同条件下均没有显著差异(扩展数据图6f–j),表明肝源性EVs对胰岛素的加工或合成没有影响。总之,这些实验表明,肝源性EVs可以增加葡萄糖氧化,并快速直接增强从小鼠分离的胰岛中的GSIS。
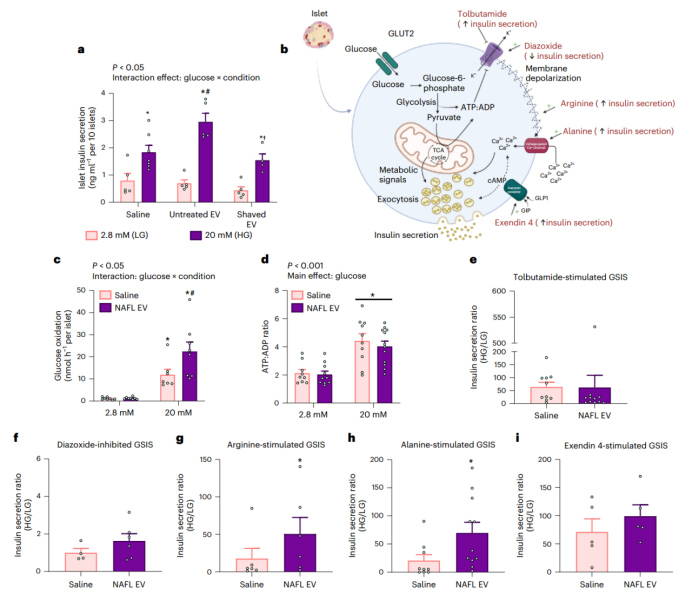
图7. 肝脏分泌的EVs直接刺激小鼠胰岛中的GSIS
扩展数据图6. 肝细胞外囊泡对胰岛素分泌和细胞胰岛素含量的影响
拓展阅读
胰岛感应葡萄糖分泌胰岛素过程
当血浆葡萄糖浓度升高时,葡萄糖会通过质膜上的葡萄糖转运蛋白(GLUT1(人)/ GLUT2(啮齿动物))进入胰腺β细胞中。之后葡萄糖会进入糖酵解途径生成丙酮酸,并经氧化磷酸化产生ATP。ATP 是β细胞中胰岛素分泌的信号分子,在细胞质 ATP 或 ATP/ADP比值升高时,K+ATP通道会关闭,导致膜去极化打开L型电压门控钙通道(VDCC),之后Ca2+内流使细胞质内Ca2+浓度升高,加快胰岛素胞吐进程,从而增加胰岛素的分泌。此外,GLP-1能作用于GLP-1受体从而增强葡萄糖依赖性的胰岛素分泌。
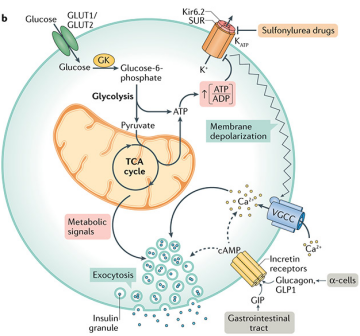
参考文献:[1] Glucose-stimulated insulin secretion: A newer perspective. J Diabetes Investig. 2013 Nov 27;4(6):511-6.[2] Campbell, Jonathan E, and Christopher B Newgard. Nature reviews. Molecular cell biology vol. 22,2 (2021): 142-158. doi:10.1038/s41580-020-00317-7
10. 人肝源性EVs改善血糖控制
为了探究了人肝脏分泌的EVs在调节血糖控制中的作用。研究人员从正在接受减肥手术的肥胖患者中提取了肝组织,制备肝脏切片,并收集和纯化EVs(图8a)。在招募的19名患者中,8名没有肝脏方面的疾病,8名患有NAFL,3名被诊断为NASH。NTA分析显示各组之间EVs分泌水平相似(图8b)。由于患者来源的EVs量较少,因此研究人员将来自具有相似组织病理学患者的EVs进行组合,并注射进小鼠用于体内分析(即2-3个患者的EVs 会被注射到一只小鼠中)。于是研究人员将生理盐水或人源的肝源性EVs腹腔注射到患有严重联合免疫缺陷(SCID,缺乏T细胞和B细胞的SCID小鼠不会排斥人类生物物质)的小鼠中 ,一个小时之后进行GTT实验发现,健康或NAFL患者的肝源性EVs能改善小鼠的血糖控制能力(图8c),并且在葡萄糖刺激条件下,EVs处理组血浆胰岛素水平会增加(图8d)。这些数据与使用小鼠来源的肝源性EVs得出的结论一致(图3a–f),均证明了肝源性EVs在高血糖期间可以改善血糖控制,并增加胰岛素分泌。

图8. 人类肝脏分泌的EVs可改善血糖控制
总结
综上所述,本研究发现了一种特殊的组织间对话模式,即肝脏EVs在高血糖时分泌,并通过增加胰腺胰岛素分泌和骨骼肌GE迅速恢复正常血糖,这也反映出了肝脏分泌的EVs在血糖控制中的重要作用。这一发现对于确定肝源性EVs诱导的信号传导,以及如何将其用于T2D的药理学干预具有重要意义。
原文链接:https://www.nature.com/articles/s42255-023-00971-z
关注微信公众号代谢学人
了解更多代谢前沿资讯

https://m.sciencenet.cn/blog-3483272-1429260.html
上一篇:代谢学人——Nature Metabolism:线粒体:我裂开了
下一篇:代谢学人-Cell :抑脂的风,还是吹到了肝脏~