博文
《自然》:栾升团队发现胞质钙稳态协调植物生长和免疫平衡
||
2024年2月29日,美国加州大学伯克利分校栾升教授课题组于Nature期刊在线发表题为 “Mechanisms of calcium homeostasis orchestrate plant growth and immunity” 的研究论文。
本研究揭示了Ca2+稳态的调节可以作为植物能在生长和防御之间进行转换的分子开关机制之一,不仅在植物钙信号转导的领域具有重要的理论价值,而且对培育新型高产抗病农作物也具有潜在的指导意义。

钙(Ca2+)不仅是植物的必需营养元素,而且是细胞信号转导中的重要第二信使。通常,植物细胞外的Ca2+浓度比游离在细胞质的Ca2+浓度高出上万倍。这种巨大的浓度梯度可以促使特定条件下细胞质Ca2+浓度迅速增加,从而作为植物用来响应环境变化的关键信号机制。然而,细胞质长时间存在过量的Ca2+对植物细胞具有毒性,并且抑制生长。因此,细胞内的钙信号通常都具有 “瞬时性” ,即需要及时地清除细胞质中过量的Ca2+以降低其毒性并重置钙稳态。一个典型的受钙信号调节的生理过程是植物免疫反应。
大量研究结果表明模式触发免疫(PTI)和效应因子触发免疫(ETI)都需要各种通道蛋白介导Ca2+流入细胞质,并通过钙信号调控下游的各种免疫应答过程。近年来,研究人员已经深入研究了植物免疫过程中的Ca2+流入细胞质(即钙信号起始)的钙通道蛋白及其激活机制。然而,人们对于Ca2+流出机制以及Ca2+稳态调节(即钙信号消退)的了解依然非常有限,并且对这些机制如何影响植物的生长和免疫尚不清楚。
这项最新研究首次报道了拟南芥中两条完整的信号传导途径,它们都汇聚于激活液泡膜上的钙氢逆向转运体(CAXs,Ca2+/H+ antiporter),在不同的生理条件下清除植物细胞质内过量的Ca2+。第一条途径发生在植物正常生长于土壤过程中,是在响应外部Ca2+水平时激活的,由定位于液泡膜的钙感受器蛋白CBL2/3以及相互作用的蛋白激酶CIPK3/9/26形成的复合体介导,它们通过磷酸化CAX1/3自抑制结构域中的一组丝氨酸簇(S-cluster)来激活CAX1/3,从而降低周围环境中的钙对植物潜在的毒害。第二条途径是发生在植物遭遇微生物入侵时,由PTI相关的免疫信号激活,涉及免疫受体复合物FLS2–BAK1以及相关的细胞质激酶BIK1和PBL1,它们通过磷酸化CAX1/3中相同的S-cluster来促进细胞质Ca2+的清除并调节免疫中的钙信号。这两条信号转导途径前者是Ca2+依赖性的(CBL–CIPK-CAX/3)而后者是钙非依赖性(FLS2–BAK1–BIK1/PBL1-CAX/3),它们通过在不同的生理条件下调节细胞质Ca2+稳态,通过激活水杨酸合成和信号途径,来实现植物生长和免疫的平衡。
环境Ca2+诱发细胞质Ca2+浓度波动
在众多植物生长发育所必需的大量矿质营养中,钙或许是一种最特殊的元素,因为细胞质内游离态的钙离子([Ca2+]cyt)浓度通常比细胞外低好几个数量级,而[Ca2+]cyt的浓度波动或维持稳态是所有钙信号转导产生的基础。为了理解植物细胞如何维持较低的[Ca2+]cyt,研究人员首先通过表达新一代钙指示荧光蛋白MatryoshCaMP6s的转基因拟南芥植株来检测外界的Ca2+浓度如何影响植物细胞内的[Ca2+]cyt。研究发现,当外部Ca2+浓度介于5-10 mM时引起了短暂的[Ca2+]cyt激增,然后迅速回到静息水平,同时在此钙浓度下植物的生长并未受到严重抑制;然而,当外部Ca2+浓度达到25 mM或更高时能够引发长时间持续的[Ca2+]cyt升高,并伴随着植物生长的停滞和叶片的坏死。同时,外界高浓度的Ca2+触发了防御反应相关基因的显著诱导表达(图1)。这些结果表明,虽然较高的外界Ca2+浓度会引起短暂的[Ca2+]cyt攀升,但只有超过一个阈值时才会发生持续[Ca2+]cyt升高,从而导致毒性、生长抑制和防御反应。由于在自然环境中,钙浓度通常在1-5mM范围内,因此对植物细胞而言,在短暂增加的[Ca2+]cyt快速反应后,Ca2+会被活性转运蛋白排出细胞质外,这也意味着在长期的环境适应中植物体内进化出快速清除细胞质内过多Ca2+的机制。
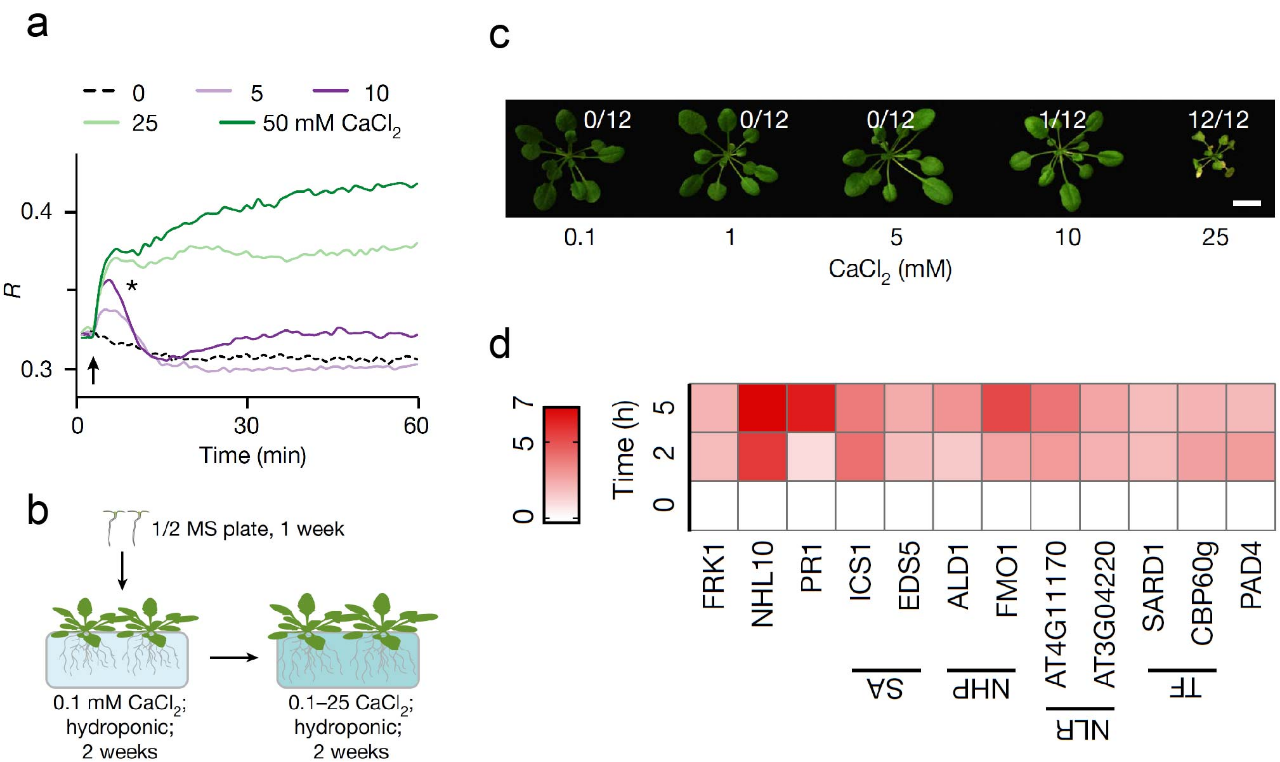
图1
Ca2+依赖的CBL-CIPK信号通路激活CAX1/3转运Ca2+
植物中主要有两类Ca2+外排转运蛋白:Ca2+-ATPase泵(ACAs)和Ca2+/H+逆向转运蛋白(CAXs)。通过对各种缺失不同ACAs和CAXs的拟南芥突变体在胞外钙处理条件下生长表型以及其[Ca2+]cyt的动态变化的系统分析,研究人员随后锁定了拟南芥中液泡膜上的CAX1和CAX3是维持[Ca2+]cyt稳态的主要贡献者,CAX1/3可以将细胞质过量的Ca2+转运到液泡内从而降低高Ca2+的毒性。紧接着,研究人员试图解析CAX1/3的激活机制,因为先前的研究表明CAX1/3转运蛋白在异源系统如酵母中表达几乎没有活性,除非去除N端一个自抑制结构域。这暗示着植物体内存在某种针对CAX1/3自抑制结构域的调节机制来激活Ca2+转运。通过对CAX1/3的N端序列分析,研究人员注意到一个独特的四个连续丝氨酸残基的簇(S-cluster),其在所有被子植物CAX1/3同源蛋白中严格保守,这暗示着 S-cluster的磷酸化可能激活CAX1/3,从而将Ca2+区域化到液泡内。共表达分析鉴定出一些与CAX1共相关的Ca2+相关蛋白激酶,其中三个高度同源的CIPK激酶(CIPK3、CIPK9和CIPK26)引起了我们的的注意,因为课题组先前的研究表明它们能够特异地与液泡膜钙感受器蛋白CBL2和CBL3发生相互作用。
通过酵母功能互补试验,研究人员成功重组了CBL2/3-CIPK3/9/26-CAX1/3的通路。体内体外磷酸化实验证明了CBL2/3-CIPK3/9/26通过磷酸化CAX1/3 N端的自抑制结构域中的S-cluster,来激活CAX1/3钙转运活性。进一步表型实验表明cbl2/3以及cipk3/9/26突变体丧失了维持[Ca2+]cyt的能力,从而抑制了正常的植物生长(图2)。
更重要的是,进一步的研究表明,钙感受器蛋白CBL2和CBL3的钙结合能力是整个系统激活所必需的。这些结果揭示了植物细胞如何通过一个精细的Ca2+依赖的“自主反馈调节”机制,响应环境信号并维持关键的细胞质Ca2+离子稳态,从而保证植物的正常生长和发育(图3)。

图2
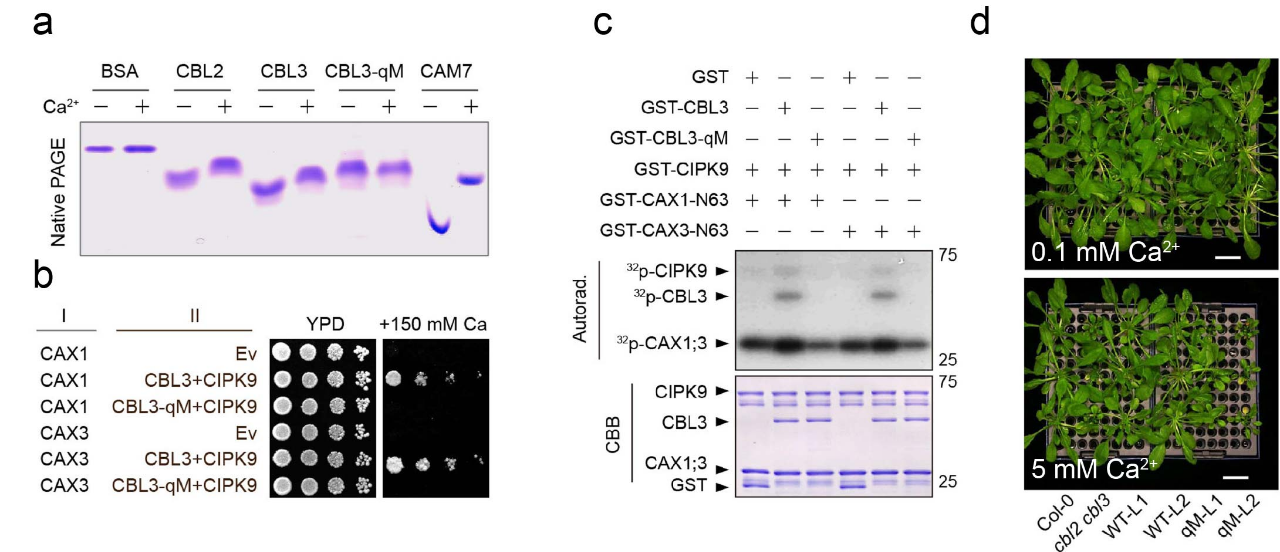
图3
模式触发免疫PTI通过免疫受体激活CAX1/3
除了应对外界环境中波动的Ca2+(如土壤中的Ca2+),植物还时刻面临着病原微生物的挑战,这导致免疫相关的[Ca2+]cyt信号出现,从而影响[Ca2+]cyt稳态。用MatryoshCaMP6s确认了PTI和ETI都会引起[Ca2+]cyt升高后,研究人员将焦点放在了PTI中的Ca2+信号上,并且试图探索PTI引发的[Ca2+]cyt升高是否能够激活类似于环境Ca2+促发的Ca2+稳态机制。研究结果表明,当用经典的PTI促发因子flg22处理植物细胞时,CAX1/3也会迅速被磷酸化。然而,令人惊讶的是,虽然flg22通路的组分,如BAK1和BIK1/PBL1对CAX1/3的磷酸化至关重要,但破坏CBL–CIPK通路并不影响flg22处理对CAX1/3的磷酸化状态。这一发现表明,在免疫反应中,有一种不依赖于Ca2+-CBL-CIPK的[Ca2+]cyt稳态维持机制。进一步的体内和体外磷酸化试验表明,PTI途径可以使用BIK1/PBL1直接磷酸化CAX1/3,从而绕过Ca2+–CBL–CIPK途径,并且有意思的是BIK1/PBL1的靶点同样是CAX1/3 N端S-cluster(简图4)。
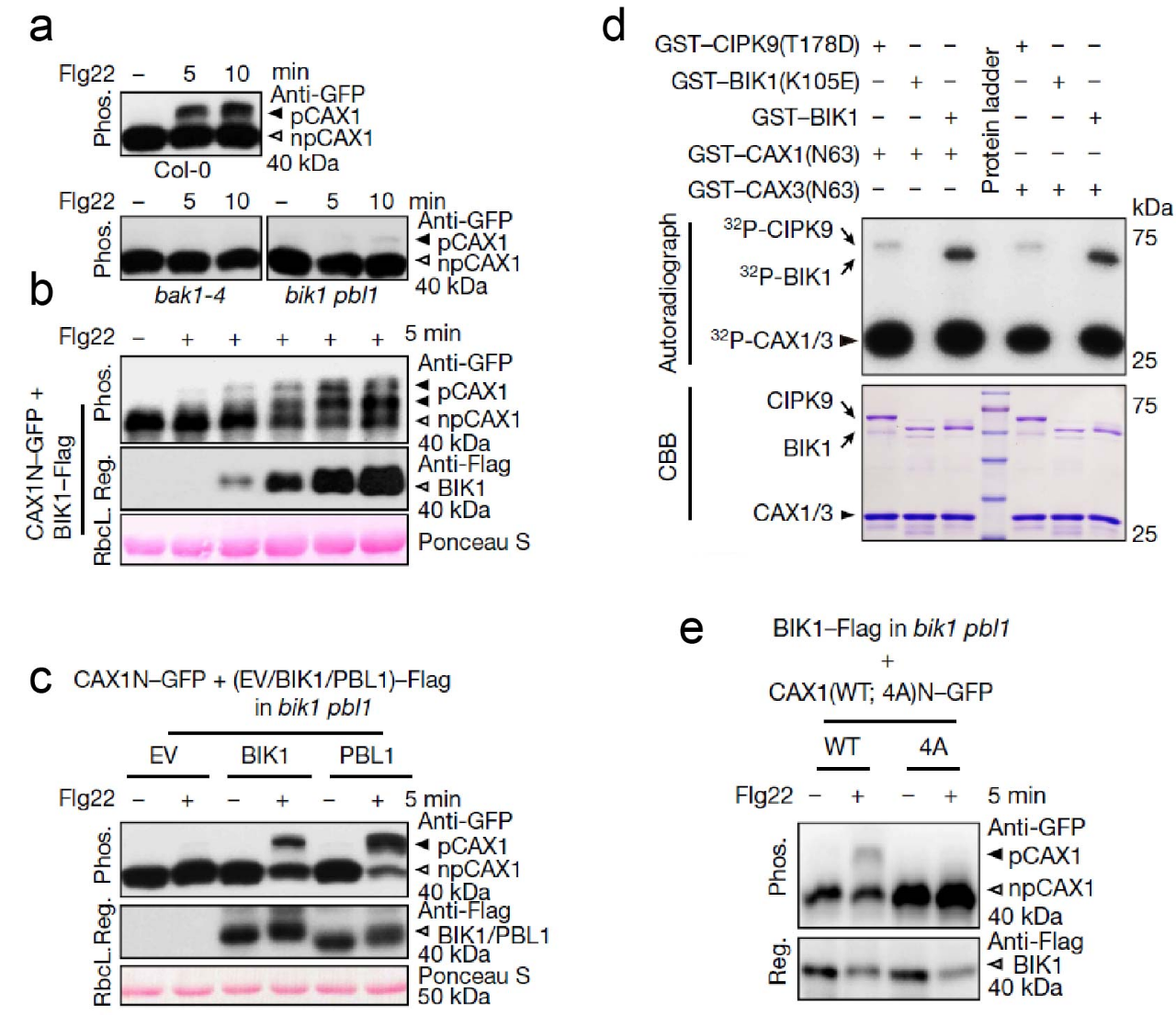
图4
非常有意思的是,从CAX1结构角度看,N端自抑制结构域阻塞了Ca2+流过CAX1的通道,而模拟CAX1磷酸化的CAX1-4D释放CAX1 N端结构域,从而解除了CAX1的抑制状态。酵母互补实验在体内也进一步证实了这一点,即CAX1-4D或者CAX3-4D能够互补高钙条件下,酵母突变体钙敏感表型(简图5)。

图5
BIK1/PBL1—CAX1/3调控Ca2+区域化进入液泡
为了研究BIK1/PBL1—CAX1/3在参与PTI产生的钙信号中的作用,研究人员设计了特异的检测Ca2+外排的试验系统来重新审视flg22诱导产生的钙信号。因为先前的所有相关研究都是观察或检测flg22处理后植物[Ca2+]cyt的净变化,而没有区分内流和外流的单独贡献。新的检测体系表明,flg22确实能够引起[Ca2+]cyt的外排,而此外排过程表现出两个特征性的波谷对应两个前后不同的阶段。与野生型植株相比,cax1/3突变体在第二阶段的Ca2+外排显著减少,表明CAX1/3的确参与了PTI过程中[Ca2+]cyt稳态维持。更重要的是,研究者观察到在PTI相关突变体fls2,bak1-4以及bik1 phl1中,Ca2+的外排类似于cax1/3突变体,即在第二阶段显著减少,这表明PTI免疫信号转导的组分不仅是Ca2+信号产生所必需的,也是Ca2+信号消退所必需的。FLS2-BAK-BIK1/PBL1信号组分通过同时作用于钙内流通道CNGC2/4以及Ca2+/H+逆向转运蛋白CAX1/3,精细调节PTI过程中“钙指纹(calcium signature)”。进一步实验表明,CAX1/3介导的Ca2+信号消退,也参与了效应子触发的免疫ETI(图6)。

图6
Ca2+失衡诱发激活水杨酸信号途径
通过以上的研究,确立了[Ca2+]cyt可作为协调植物生长和免疫的关键信号。由于cax1/3突变体缺乏细胞质Ca2+清除能力,其可作为一个良好的高[Ca2+]cyt研究模型。在正常土壤条件下,cax1/3突变植物即表现出生长受阻,显示出[Ca2+]cyt持续升高、水杨酸产生增加以及防御相关基因的组成型高表达,与野生型植物受到病原菌入侵后的反应相似。当cax1/3突变植物在低Ca2+(0.1 mM)环境中生长时,这些典型的“自免疫”表型消失,而在转移到较高Ca2+(10 mM)条件两天后又重新出现。进一步研究表明,cax1/3突变体中的Ca2+依赖性自免疫激活和生长抑制表型可能与水杨酸的信号传导途径有关,因为水杨酸合成(sid2-2)缺陷,水杨酸受体(npr1-1 npr4-4D)突变,或水杨酸信号通路正调节因子(eds1-2)的缺失都能有效的恢复cax1/3突变体的生长抑制和自免疫的表型。然而,这些高阶突变体的[Ca2+]cyt维持能力,与cax1/3突变体相当,表明水杨酸合成和信号传导途径并不影响[Ca2+]cyt的稳态,而是在高[Ca2+]cyt的下游起作用,从而介导生长抑制和免疫反应(图7)。
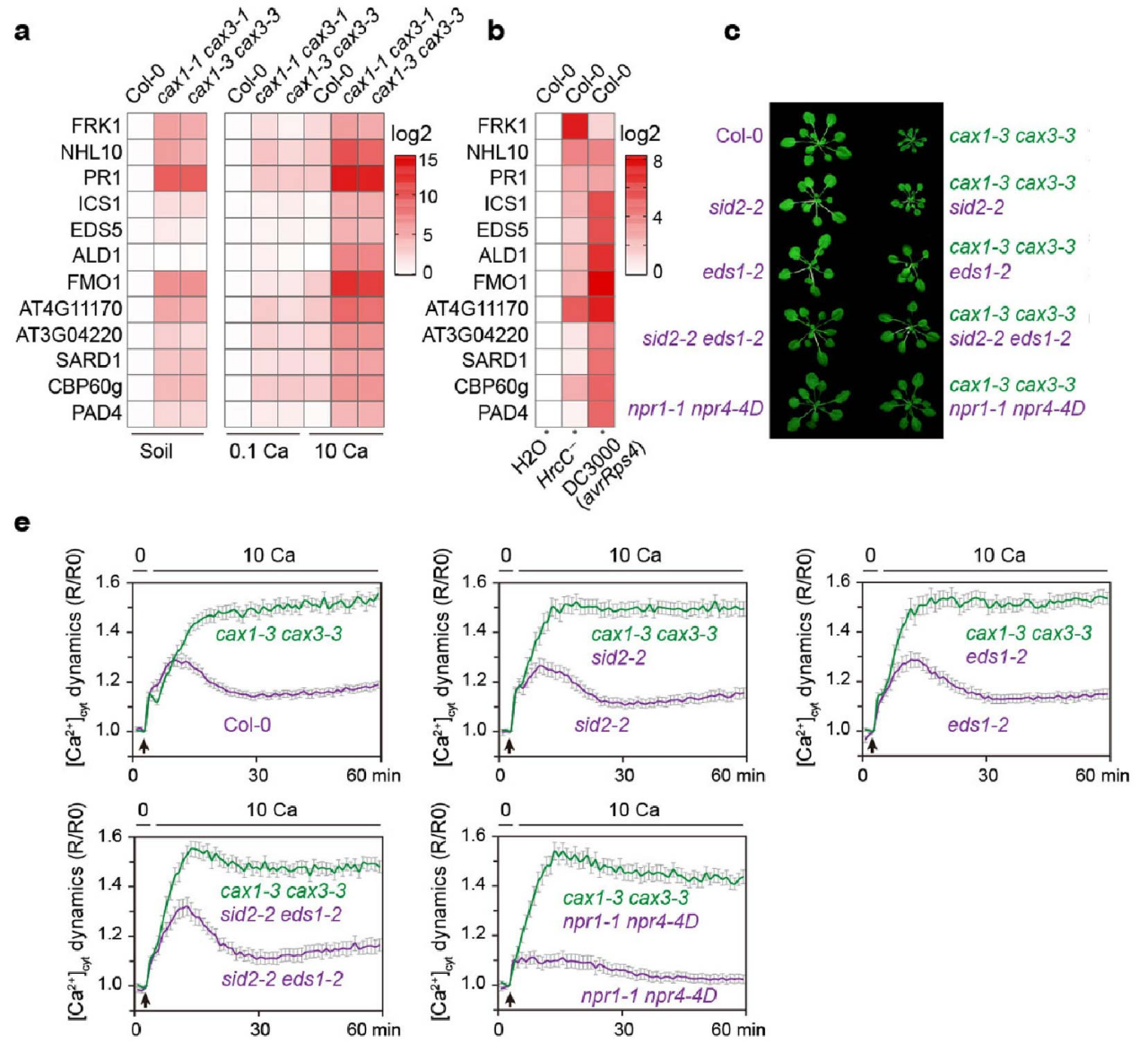
图7
无论生长还是防御都是植物在其生命周期中需要做出的最常见抉择。本研究揭示了Ca2+稳态的调节可以作为植物能在生长和防御之间进行转换的分子开关机制之一。在正常土壤生长条件下,植物通过Ca2+–CBL–CIPK–CAX1/3通路将Ca2+区域化到液泡内,以维持[Ca2+]cyt的静息态水平。这条途径代表了一种依赖Ca2+反馈循环的自主调控机制以适应自然环境中的钙水平。此外,在PTI的免疫反应中,FLS2—BAK1—BIK1/PBL1通路同样激活了CAX1/3,从而塑造了免疫中的钙信号。这其中,BIK1/PBL1对钙信号具有双重功能:一方面磷酸化激活CNGC2/4通道起始钙信号;另一方面,磷酸化激活CAX1/3转运蛋白解除钙信号。CAX1/3的破坏会导致持续的[Ca2+]cyt升高,从而触发水杨酸依赖的免疫并导致生长抑制。这项研究不仅在植物钙信号转导的领域具有重要的理论价值,而且对培育新型高产抗病农作物也具有潜在的指导意义。栾升教授领导的课题组多年来致力于植物钙信号产生和解码的分子机制研究(Wang & Luan, 2024; Luan & Wang, 2021),在该领域有着丰硕的研究成果。例如,课题组多年前就证明了CNGC2和CNGC4组成的钙通道复合体是介导植物PTI过程中钙信号产生的关键组分之一(Tian et al,2019)。近年来,课题组还成功鉴定出植物新型的钙通道MLO家族,并阐释了其中成员在植物生殖过程中的重要功能和激活机制(Gao et al, 2022; Gao et al., 2023)。在植物钙信号解码方面,栾升教授在二十多年前首次报道了植物中特异的Ca2+感受器CBL蛋白极其下游的激酶CIPK,并多年来深耕于这两个蛋白家族的功能鉴定和分子机制,受到了国内外同行的广泛关注和跟踪研究,到今天CBL-CIPK系统仍然是植物钙信号与逆境相关研究的热门课题之一(Tang R-J et al, 2020)。最新发表的这篇研究论文凝集了栾升教授领导的团队多年来在该领域系统性研究的成果,从细胞、分子、生化和遗传各个水平展示了钙转运蛋白通过钙依赖和钙不依赖两条不同途径的调控方式,从而阐释了钙稳态调节作为植物生长和防御转换的分子开关机制。课题组中的王超博士,唐仁杰博士为该论文的共同第一作者;栾升教授为通讯作者;斯坦福大学的徐晓庶博士和加州大学伯克利分校的卢毅博士, 以及访学生寇森浩,本科生Kenda Rauscher、Angela Voelker为该课题的共同作者。该项目受到美国NIH和NSF项目资助,同时感谢众多课题组同行无私的分享钙转运和免疫相关植物突变体和其它实验材料。
相关论文信息:
DOI:10.1038/s41586-024-07100-0
https://m.sciencenet.cn/blog-3423233-1423524.html
上一篇:《癌症研究》:郗乔然/张力伟合作阐述吉西他滨在H3.3K27M突变型弥漫内生型脑桥胶质瘤中的作用机制
下一篇:《自然》:王凌华/Humam Kadara合作团队利用单细胞和空间转录组揭示肺腺癌的细胞起源