博文
光遗传学
 精选
精选
||||
人们常把人脑与电脑相比,两者都是由大量相似的基本元件构成:计算机里面有数亿个晶体管,大脑里有数百亿个神经细胞(图1)。在计算机和大脑中,基本元件之间都互相连线构成巨大的网络,在网络中利用电信号快速传递着海量信息。神经科学家怎样通过测量大脑中的信息流来了解大脑的功能呢?传统的方法是用用电极测量神经细胞上的电信号,像电话窃听器一样来了解他们是怎样工作的。这个办法也叫“电生理”,虽然已经在历史上延用百年,但缺点是很明显的:在一个含有数亿个神经细胞互动的过程中只测量其中几个神经细胞活动,就好像试图通过电视屏幕上几个像素的闪烁来猜测一个电视剧的内容。
早在1960年代,耶鲁大学年轻的科恩教授(Lawrence B. Cohen)就开始想办法引进一个光的元素,把神经电信号变成光,让神经科学家看到大脑活动的“电视剧”。他的想法是利用一种化学染料把神经细胞染色。染料的分子并不进入神经细胞,而只附着在细胞膜上。这样当神经细胞产生电信号时,染料的分子结构受到电场影响而改变颜色。这样就可以通过测量染料的荧光来对神经组织进行成像。这种“电压敏感染料成像”方法转眼做得风声水起,在顶级杂志上文章一篇接一篇;他的学生找工作时只要放一小段震撼人心的“脑电影”就能轻易俘获满堂听众的芳心,顺利地得到工作。但是这种方法也有个明显的缺点,就是对神经细胞没有选择性,不同神经细胞对光信号贡献没法分开。我们知道大脑中有兴奋和抑制两种神经细胞,就像汽车的油门和刹车,兴奋的负责“嗨”,抑制的负责瞌睡。如果有一种方法能分清楚这两种不同细胞的信号,那研究工作就更上了一层楼。可是化学染料对神经细胞的选择性是很差的,远远做不到分辨不同来自细胞的信号。这个问题整整为难了一代神经科学家。
科恩小组的工作主要是利用大学长长的暑假,把实验室搬到在麻省海边,位于鳕鱼角的伍滋霍尔海洋生物站(WoodsHole Marine biological station)。那几十年是美国科研的黄金时代,在伍滋霍尔聚集了一大批心怀各种梦想的神经科学家。他们白天躲在各自的实验室里工作,晚上则聚在海滩或镇上的小酒吧里高谈阔论。谁有个新发现身边马上聚满粉丝,新闻转瞬传遍世界,根本等不到文章发表。科恩的学生乘着电压敏感染料的东风发明了另一种染料,它可以进入细胞并随着细胞内钙离子浓度的变化发光。钙离子在一切细胞中都有非常重要的作用,能做“钙成像”的消息一下引爆伍滋霍尔,他火得连在上厕所的时候,站在旁边尿尿的人都会突然提个问题。
在这群伍滋霍尔科学家中有个谦和的日本科学家OsamuShimomura(下村修),成天拿着捞网在海边收集水母。伍滋霍尔海水温暖,夏天傍晚阵阵清风掀起涟绮,海面成千上万的水母会竞相发出绿色荧光,美如梦幻。下村多年来一直着迷于这种发光的现象,并于1960年代在普林斯顿大学纯化了这种的绿色荧光蛋白。年复一年,伍滋霍尔夏末的学术讨论会上,他在做报告时让把灯全关掉,在黑暗中他从口袋里拿出两只试管,当溶液混合时会产生幽幽的鬼火似的蓝光。报告厅里科恩照例坐在角落里,瞪着一双牛眼不知道在想什么。下村的荧光蛋白与科恩的光学成像多次在报告厅里相遇,但从来没擦出火花。因为分子遗传学的大潮还没有涌现,光学遗传学的弄潮儿还站在海滩上呢。
1990 年代,下村的水母荧光蛋白迅速成为光学遗传学领域中的第一个明星。首先,它的基因被克隆,并可以在其他物种中表达。由此只要把它的基因转移到某些细胞中,这些细胞就可以在组织中发亮,给科学家指引目标。1995年,加州大学圣地亚哥分校的钱永健发现改变蛋白分子上一个氨基酸可以时其发光大大加强而且更稳定。那几年有个“荧光蛋白热”,几个小组使用各种方法改变绿色荧光蛋白,不但使其更亮更稳定,而且也产生出红,蓝,黄,青等不同颜色的蛋白。这项工作终于在2008年与下村修和Martin Chalfie一起获得了诺贝尔化学奖。
如果用转基因的方法让脑细胞随机地表达几种不同颜色的荧光蛋白,在紫外光下每个脑细胞就会出五彩斑斓的荧光,把每个细胞看得清清楚楚(图一)。这项技术也叫“脑彩虹”(Brainbow)[1]。

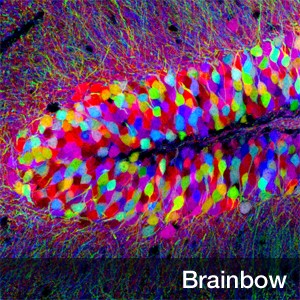
图1.Brainbow(脑彩虹)项目获得的荧光标记海马皮层细胞图和荧光标记视网膜方向选择性神经元图。左图展示的是使用脑彩虹技术将不同的海马皮层细胞用不同颜色区分开来。图中彩色大斑块就是细胞胞体,纤细的须状物就是神经树突。左图作者:Tamily Weissman, Harvard University右图是视网膜中参与方向信息识别的神经节细胞。树丛一样的是神经节细胞的树突,一根根长长的线是轴突,轴突都汇集到眼球的视盘(右图下部)然后到达大脑的视觉中枢。右图作者:JoshR. Sanes, Harvard University。图片来源:http://www.cell.com/pictureshow/brainbow
有了可以通过遗传学手段标记细胞的荧光蛋白,光学遗传学下一步的目标就是怎样利用荧光蛋白显示神经细胞的活动,以及怎样利用光来指挥和控制神经细胞的活动。
光学遗传学方法测量神经细胞的电活动
一个神经细胞的活动有两个重要的指标:第一个是细胞膜电位的变化,第二个是细胞内钙离子浓度变化。下面分别描述测量这两个指标的方法。
神经细胞的膜电位变化是神经细胞的活动最基本的信号,当膜电位变化时,细胞膜上镶嵌的许多蛋白质分子都会改变形状,因此,改变膜电位是一个细胞指挥自己身上亿万个蛋白分子统一行动的信号。这类随膜电位改变形状的蛋白分子也叫电压敏感蛋白。如果用基因工程的办法,把电压敏感蛋白和荧光蛋白连接起来,当膜电位改变时,电压敏感蛋白的改变就会影响荧光蛋白的结构,从而改变了后者的发光特性。这样就可以利用荧光来看神经细胞膜的膜电位变化了。

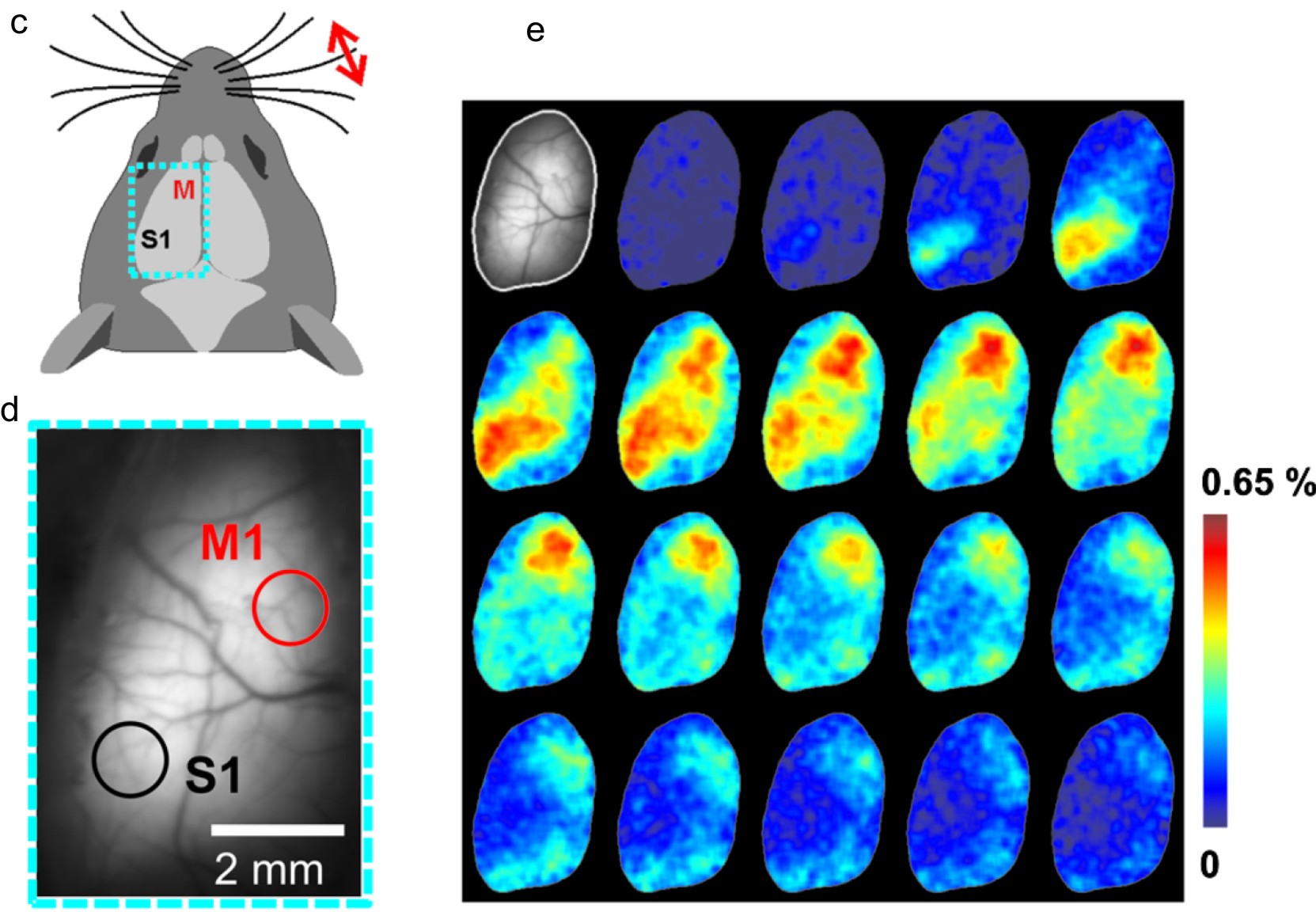
图2. 膜电位变化指示蛋白(a)ASAP1[2]的设计基于环段置换绿色荧光蛋白(circularly permutedgreen fluorescent protein,cpGFP)。当膜内电位由负变正去极化,插入膜中的结构域被推斥向膜外,cpGFP结构变化,被蓝光激发的绿色荧光量下降。(b)一部分神经元表达电位指示蛋白,当细胞兴奋时,荧光强度下降。(c)小鼠大脑皮层示意图(d)暴露的大脑表面照片(e)电位变化成像图,在刺激后每隔20毫秒拍一张荧光图片。(c, d, e图来源于Akemann W 的综述[3])
图2a举例说明这种思路。图2a左面的绿色桶形是绿色荧光蛋白分子,灰色的长方块是电压敏感蛋白,它的分子在细胞膜(黄色)的内外穿插四次。当神经细胞不活动时细胞膜处于内负外正的状态,这时电压敏感蛋白整齐排列,当荧光蛋白分子受到蓝光(蓝箭头)的照射时会发出绿色的荧光(绿箭头)。当神经细胞兴奋时膜电位会改变,成为内正外负(图2a右)这时电压敏感蛋白分子形状发生改变,牵扯到荧光蛋白也改变形状,造成绿色荧光大大下降。如图2b所示,当用转基因的办法让神经细胞携带这种荧光蛋白-电压敏感蛋白复合体时,在大脑皮层不活动时可以看到较强的荧光(图2b,左)而当大脑皮层活动时荧光就会减弱(图2b右)。
图2c-e显示这种光遗传学方法测量神经电活动的一个应用实例。小鼠的胡须是其非常重要的感受器,当一根胡须被拨动时大脑皮层感觉区(S1)会有很强烈的神经活动。为了显示这种当对小鼠的大脑皮层进行荧光成像时(图2c中的蓝方框,放大到图2d中),当一根胡须受到拨动时就会看到皮层不同的区域荧光顺序地减弱,表示神经活动在皮层上扩布(图2e)。
光学遗传方法测量细胞内钙离子
钙离子参与了细胞内很多生理过程,如细胞内信息传递,基因表达,神经递质释放等等。细胞内的钙离子浓度是非常低的,胞外钙浓度是胞内的十万倍以上。虽然有这个巨大的浓度梯度,但是钙离子不能透过细胞膜,必须通过特殊的钙离子通道(图3a中绿色的蛋白分子)才能进入细胞。神经细胞在静息的时候钙离子通道都是关闭的。而当膜电位变化的时候对电压敏感的钙离子通道会大批开放,让胞外的钙离子就会迅速涌进胞内,造成一个突然的钙高峰。那么如何来观察这个细胞内钙浓度的突然增加呢?生物基因工程学家的思路与设计前述的电压敏感荧光蛋白类似,将能与钙离子结合的蛋白组合CaM(钙调蛋白)和它的亚基M13分别连在绿色荧光蛋白的两个位置上。当这个融合蛋白不结合钙离子的时候,绿色荧光蛋白不会发出荧光。而当CaM和M13结合了钙离子,这两个蛋白就会更加紧密的靠在一起,绿色荧光蛋白的结构完整的合起来,就发出了明亮的荧光(图3b)。基因工程重组钙敏感荧光蛋白就是通过这个方法来构建的。
图3c-d显示这种光遗传学方法测量神经细胞内钙浓度变化的一个应用实例。表达有钙敏感荧光蛋白的神经细胞在不兴奋的时候有比较弱的荧光,而当该神经细胞兴奋时,细胞外的钙离子进入细胞内,细胞内的钙敏感荧光蛋白结合了流入的钙离子之后,荧光强度变亮,细胞发光,观察者就知道这个细胞参与了此时此刻的神经信息处理过程。

图3.钙浓度变化指示蛋白示意图(a)钙离子参与细胞活动的方方面面.钙离子是细胞信号传导中的重要信使,因为一旦它们进入细胞质,钙离子会结合并激活很多酶和蛋白质。由下图所示的细胞为例,钙离子通过AMPA受体,NMDA受体等离子通道进入细胞,可引发一系列突触后膜的活动,可能引发突触结构性的变化,比如更多的AMPA受体插入膜上。在细胞胞体内流的钙离子,或者细胞内钙库释放的钙离子结合钙调蛋白,可影响基因表达。在神经细胞轴突末梢流入的钙离子,是神经递质释放的前提条件。(b)基因工程重组钙敏感荧光蛋白(genetically encoded calcium indicator由Robert E. Campbel设计。(c)细胞兴奋前后荧光量的变化 (d)荧光强度随着细胞内钙的浓度变化而波动[4]
用光控制神经细胞的活动
在经典的神经生物学实验中,激活一个神经细胞大致有两种方法,物理的和化学的。物理的方法就是电刺激,例如将金属电极放在神经细胞旁边,改变细胞外电场,从而激活细胞。化学的方法,就是施加神经递质类的小分子,或者作用于细胞受体的药物。而光遗传学提供了一个全新的方法,就是直接用光来准时刻准位点的刺激细胞。光遗传学是怎么做到的呢?
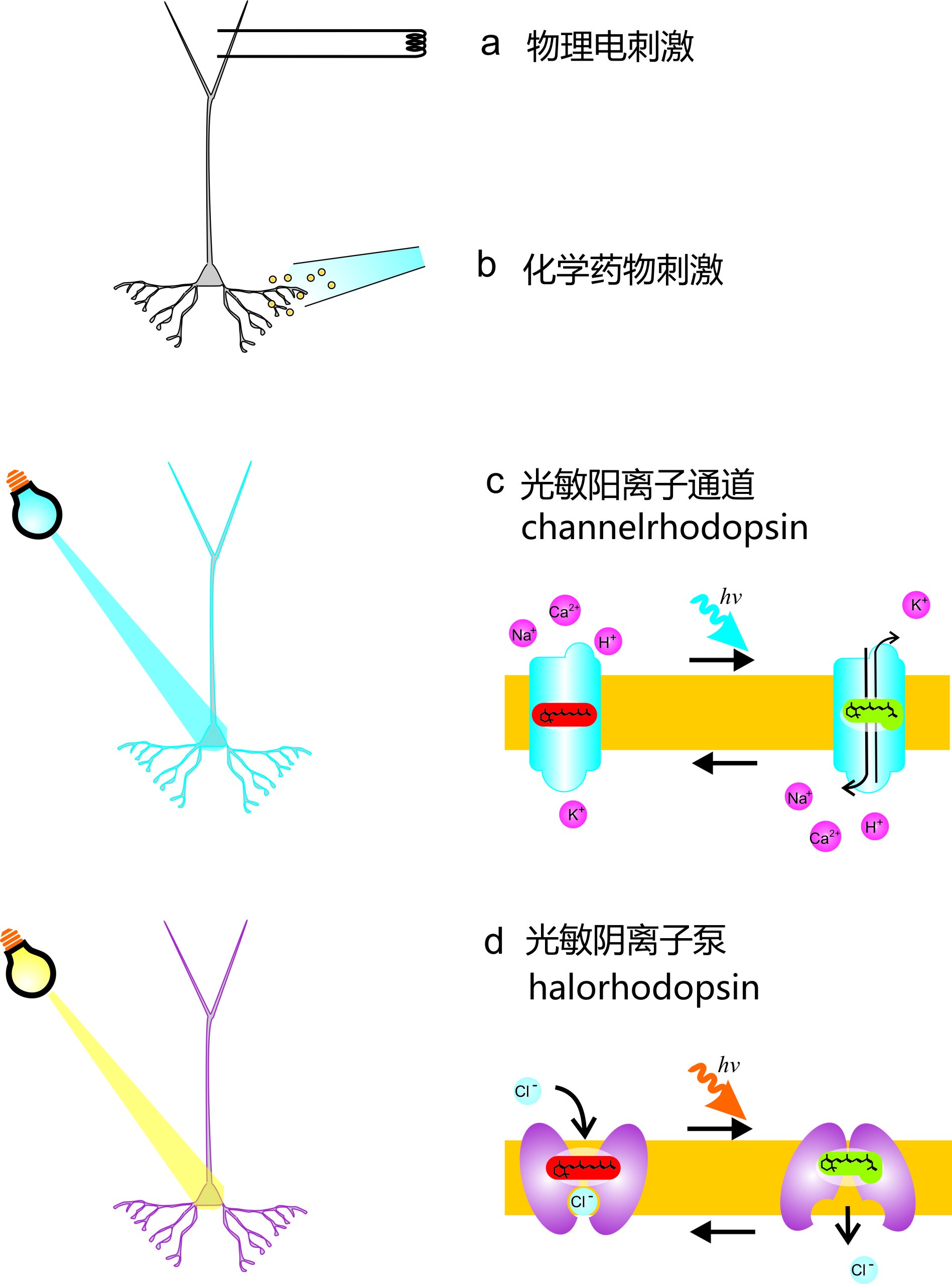
大家知道人类的眼睛就是一个光信号-生物电的光电转换系统,那么是否可以用眼睛的方法来进行光电转换呢?让我们首先用一张图来大致了解人类眼睛的光信号转换为细胞电信号的过程[图4]。感兴趣的朋友可以仔细阅读图4的图解,但是我们其实只需要瞄一下这张图有个直观感受,在感光细胞中,要将光信号转换为使细胞电生理兴奋状态的改变,需要十几个蛋白协同工作;但是光遗传学中,只需要一个光敏蛋白(例如光敏通道channelrhodopsin,ChR2),就可以使细胞兴奋。这在科研应用中非常重要,因为在一个细胞中多表达几个蛋白,比只表达一个蛋白的难度系数可是成几何级数增长的。而细胞本身也无法承载那么多外源蛋白的额外负担,而且这么多外源蛋白的表达程度还必须保持相互平衡状态,这简直是不可能的任务。甚至可以说,光敏通道这一种蛋白,就几乎可以代替整个感光细胞这一精密复杂的光电转换生物系统,作为感光受体(photoreceptor)。

图4. 感光细胞光电信号转换过程简化说明以及光敏通道示意图:(a)光强不变的时候,感光细胞中的有高浓度的cGMP,cGMP门控阳离子通道(cGMP-gated channel)开放,细胞一直处于去极化,释放递质谷氨酸(glutamate,Glu)的状态。在光强增加的时候,光子被视紫红质(rhodopsin,Rh)吸收。视紫红质由吸收光子的小分子视黄醛(11-cis-retinal)和视蛋白(opsin)组成。视紫红质其实是一个G蛋白偶联受体,它激活了G蛋白(transducin)。transducin释放GDP,结合GTP。Transducin的α亚基与βγ二聚体分离,α亚基结合并激活磷酸二酯酶(phosphodiesterase,PDE),PDE水解cGMP成GMP,cGMP浓度下降,cGMP门控阳离子通道关闭,细胞膜电位下降超极化,电压门控钙通道关闭,递质Glu释放减少。值得一提的是,感光细胞内还有很多精细调控和反馈机制,还有很多蛋白并未在图中提到。而没有这一大群蛋白,感光细胞的功能是无法正常发挥的。(b)以光敏通道ChR2为例,光敏通道只需要一种蛋白即可,当与ChR2共价结合的all-trans-retinal吸收了光子,促使ChR2分子构象发生变化,蛋白中间出现阳离子通道,Na离子和Ca离子的内流就可使细胞内电位上升,细胞去极化兴奋,如果表达ChR2的是神经细胞,膜电位去极化可激活在轴突末端的电压门控钙通道,轴突末端的钙内流可促使神经细胞释放神经递质囊泡。
2002年Gero Miesenböck实验室[5]首先尝试了这一个大胆的设计,他把视紫红质表达在肌肉细胞上,再把果蝇砍去头,这样果蝇的扇翅膀的运动就不能被自身的运动神经所控制。然后,再把光打在果蝇身体上,没头的果蝇竟然拍翅膀了!这是实现光控细胞活动最早的一个成功的实验。
因此当2003年左右当PeterHegemann 实验室连续发变了两篇微生物中的单个光敏离子通道(channelrhodopsin)ChR1[6]和ChR2[7]时,许多神经生物学家精神为之一振,世界各地同时有四个实验室尝试表达于哺乳动物细胞,可谓英雄所见略同。他们有美国斯坦福大学的KarlDeisseroth实验室[8],美国Case Western Reserve University 的Lynn T. Landmesser 和 Stefan Herlitze实验室[9],日本的Hiromu Yawo实验室[10],以及美国Wayne state University 的潘卓华实验室[11]。Karl Deisseroth实验室的文章是最先发表于2005年8月Nature Neuroscience,但是只做了体外培养的细胞表达,值得一提的是发展了CRSIPR/cas9工具的张峰也参与了这篇工作,他是二作。仅仅迟了两个月的Lynn T. Landmesser 和 Stefan Herlitze实验室就做得全面很多,有体外培养海马皮层神经元表达,以及在体脊髓神经元表达。日本的HiromuYawo实验室比Deisseroth迟了3个月,但是也做了体外培养细胞系PC12的表达和改变细胞兴奋性的实验。但是最晚于2006年4月发表的潘卓华老师实验室的工作做的最全面,从体外细胞表达到小鼠在体神经细胞表达和电生理记录一应俱全。实际上据潘卓华老师介绍,在2004年底他的在小鼠视网膜体内表达的实验就已经完成,而且依次投递了Nature和Science,但是都未被接受,甚为遗憾。这部分历史在Peter Hegemann的综述[12]中有客观叙述。
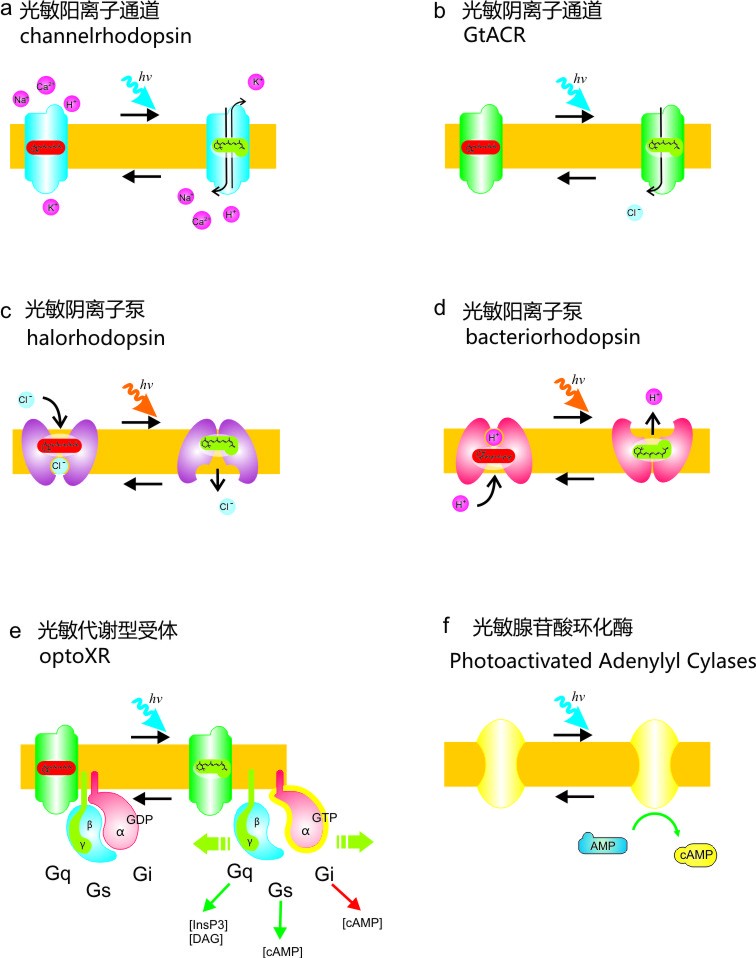
图5光敏蛋白工具箱(a)光敏阳离子通道(b)光敏阴离子通道(c)光敏氯离子泵 (d)光敏氢离子泵(e)光敏代谢型受体(f)光敏腺苷酸环化酶
2006年后,光遗传学的时代来临了,光遗传学给物理光学,光化学和生物学提供了新的结合点。结合了基因工程技术和最前沿的光学技术。这个方法有很多优点,例如在体表达的实际操作性高,准确时效性,减少创伤,无异物侵入组织等等。可以用定位的光纤来局部刺激细胞,也可以设计弥散光来大范围的刺激脑区。许多实验室开始广泛参与光遗传学的研究工作,有些实验室使用光敏通道来代替经典电极刺激,有些实验室寻找更多的光敏蛋白来丰富这一工具箱,有些实验室希望利用光敏蛋白来提供临床治疗应用。
经典的基因工程研究思路中,曾经是把来源于不同物种的蛋白在动物模型上表达,或者将原始蛋白和荧光蛋白形成融合蛋白等等。但是光敏通道的发现似乎一下子开拓了科研人员的想象。从光敏蛋白ChR2的结构来看,完全不像是个经典的通道结构,而像一个代谢型G蛋白偶联受体,ChR2是一个7次alpha螺旋跨膜结构,但是竟然在7个螺旋中有离子的通道。因此研究者脑洞大开,认为理论上可以通过基因工程的改造,有可能将所有原本不受光子激活的蛋白都改造为对光敏感。就像在2002年发现的如图5f的光敏腺苷酸环化酶[13](Adenyl cyclases)一样。使用这个思路,2006年Ehud Y Isacoff 和Dirk Trauner实验室合作,成功构建了光敏谷氨酸离子通道(LiGluR)[14]。Chandra L Tucker实验室[15]和Wendell A. Lim实验室[16]在2009年左右分别独立的设计了光控制的蛋白相互作用和光控制基因转录方法。
在图5中举了几个光敏蛋白的例子。如果想要使用光来兴奋细胞,可以使用图5a的光敏阳离子通道,施加蓝光可以使阳离子通过该通道进入细胞,使细胞内膜电位成为内正外负,细胞就可以兴奋。
如果想要使用光来抑制细胞活动,可以使用图5b的光敏阳离子通道、图5c的光敏氯离子泵和图5d的光敏氢离子泵。图5b中施加蓝光可以使阴离子通过该通道进入细胞,使细胞内膜电位成为内负外正,细胞就可以被抑制。图5c和5d中施加黄光可以使阴离子通过该离子泵进入细胞或者阳离子离开细胞,使细胞内膜电位成为内负外正,细胞就可以被抑制。
图5e的光敏代谢型受体,不是通过直接的把阴阳离子跨膜运动来改变细胞的活动,而是较为间接的通过光控制细胞内的化学作用来达到影响细胞的目的。
在特定细胞环路上表达光敏通道,再使用可拍摄深层组织的双光子显微镜系统,使在体观察动物神经系统活动成为可能。这也是最近十来年最大的突破。神经科学家对神经系统的认知早已从递质、激素和受体决定一切,转变为认为大脑是一个高效细胞环路。生物工程最前沿的目标应当是结合光遗传学、光学成像和组织细胞工程来人工构建细胞环路。
临床医疗研究者也在近十年尝试利用非侵入性的光遗传学手段来治疗各种疾病,例如嗜睡症[17],抑郁症[18],恐惧[19],焦虑[20],疼痛[21],帕金森综合征和失明。这些尝试给“光疗法”赋予了全新的意义。但是将感光蛋白引入人体还是需要非常谨慎的,毕竟我们对人体还不能说是完全的了解和掌握。特别是很多精神疾病涉及的脑区很广泛,使用光遗传学治疗很难刺激如此大的范围。因此,只有帕金森症以及失明的治疗在现阶段看来比较可行,因为所涉及的组织区域已明确而且是一个限定的区域。美国药监局FDA2015年已经批准了光遗传学治疗失明的临床试验。
光遗传学的开拓的全新领域也激发了科学家更多的想法:比如为什么光控受体一定要是蛋白质呢?是否可以有DNA、RNA的光感受器来加入光遗传学的工具箱?例如Richard H. Kramer实验室设计了AAQ[22]和DENAQ[23]感光小分子并用于尝试治疗失明。比如声遗传学,就是利用声波或者超声波来影响细胞;比如磁遗传学,就是利用外加磁场来无创控制细胞。未来是不是还有机械遗传学,用纳米颗粒甚至智能小机器人来提供局部提供机械刺激给细胞呢?在这个时代,anythingis possible。
参考文献:
[12]Hegemann P. Algalsensory photoreceptors. Annu Rev Plant Biol, 2008, 59: 167-89
https://m.sciencenet.cn/blog-5525-975556.html
上一篇:受“圣人婊”启发,想起BITCH这个词
下一篇:徐晓老师:图来了