博文
代谢学人--好看的生化书:三羧酸循环篇
||
撰文 | 陈俊桐 郭文秀
编辑 | 孟美瑶

引言
书接上回
上回书说道:
一分子葡萄糖被细胞摄取,
先是借助转运蛋白跨过细胞膜;
后又历经胞质糖酵解的十重关卡,
才最终“修成正果”——丙酮酸。

那么葡萄糖的闯关之路如此曲折
“新生儿”--丙酮酸肩负产能的重任
接下来要如何打拼出一番事业
为机体做出贡献呢?
一起来瞧瞧糖酵解的后续↓↓↓↓↓↓
经济又高效的有氧供能途径---
三羧酸循环(tricarboxylic acid cycle,TCA)
让我们以丙酮酸的视角在TCA循环中走上一圈
各位观众老爷们,这里先给您报个幕~

前言
如果你热爱健身,想必一定听说过肌酸(Creatine),许多厂商将其制作成保健产品,有补充能量,肌肉增力等优点,那么肌酸究竟好在哪里呢?


肌酸产品广告
肌酸在体内能够被磷酸化为磷酸肌酸(Phosphocreatine,PC),PC充当能量的储剂,在机体需要能量时能够快速合成腺苷三磷酸(Adenosine triphosphate,ATP),因此肌酸可以作为增强肌肉爆发力和耐力的补品。每天我们都要靠大量的ATP来供能,维持我们生龙活虎的状态,ATP结构中高能磷酸键的发现让”能量“不再是一个看不见摸不着的东西。
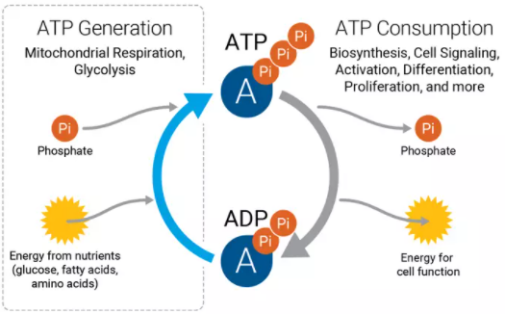
ATP的产生与消耗
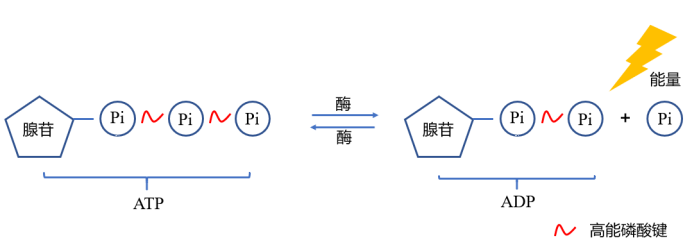
ATP分子断裂高能磷酸键为机体供能
那么机体如何获取ATP呢?上回书中(好看的生化书——糖酵解篇、好看的生化书——糖酵解篇 完结篇)我们发现一分子的葡萄糖经历了细胞质中糖酵解作用(EMP途径),消耗2分子ATP,产生4分子ATP,净剩2分子ATP,同时生成2分子的丙酮酸和NADH。
通过EMP途径,一分子葡萄糖最终只产生了2分子ATP,实在不够用啊!真正的物尽其用,可不是随便说说而已,含有6个碳原子的葡萄糖,其价值远不止于此,若机体只靠糖酵解作用来获得能量,我们可能马上就会濒临死亡。因为多细胞生物不仅要维持细胞的生长分裂,同时还要负责整个机体的新陈代谢。对我们来说,光靠EMP来产能远远不够。

因此,在氧气的存在下,细胞利用线粒体作为产能车间,将葡萄糖的酵解产物丙酮酸通过三羧酸循环(tricarboxylic acid cycle,TCA)彻底氧化分解,充分发挥产能效应。
在谈具体的TCA反应流程前,先让我们看看丙酮酸面临的去向与抉择:
丙酮酸的命运抉择--无氧还是有氧?

葡萄糖的两种利用形式
无氧呼吸
葡萄糖经历EMP作用,将NAD+转变为NADH。NAD+是氧化还原反应中传递电子的辅酶,为细胞生存过程所必要。在无氧或是缺氧状态下,细胞通过无氧呼吸将丙酮酸转变成不完全代谢产物乙醇(植物中)或是乳酸(动物中):丙酮酸在丙酮酸脱羧酶催化下,生成乙醛,进而被还原成乙醇;或是在乳酸脱氢酶的催化下,被还原成乳酸(见下图两条红色支路)。
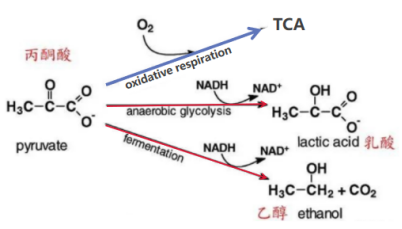
丙酮酸的去向
图片来源:(全网最科学谈酒-在喝酒与化学之间 - 知乎)
无氧呼吸只能够为机体提供2分子的ATP来维持机体基本的生存。但是,若细胞需要能量的快速供给时,无氧呼吸所介导的有机物不完全氧化过程产生的能量较少,细胞进而会加速糖酵解速率来补偿低的ATP产额,这一过程造成不完全氧化产物的积累,对细胞产生毒性(如动物的疾跑会造成乳酸堆积,破坏肌肉微细结构,导致肌肉酸痛的现象);此外,无氧呼吸也加速了对糖的消耗,有耗尽可供呼吸物质的危险 [1]。所以无氧呼吸不能满足我们的生存需要,还可能给机体造成伤害。

有氧呼吸
工作学习、交流聚餐、唱歌跳舞、看书品茶,我们的生活行为如此复杂且丰富,糖酵解或无氧呼吸的产能效率显然不能满足我们的各种需求。在氧气的参与下,瞬间豁然开朗,胞质中的丙酮酸会优先进入到线粒体内(即上图第一条蓝色支路),先转换成高能分子乙酰CoA,再经过TCA循环彻底地氧化分解成CO2和H2O。TCA循环能够合成大量的ATP分子,为生命体的各种生理过程(如肌肉收缩、生物合成、细胞运动、细胞分裂、主动转运、神经转导等[2])提供能量;同时,TCA循环这个能量圈圈,除了作为细胞能量这一“货币生产厂”以外,还是“底物中转站”和“代谢交流所”,可谓是多能代谢途径于一圈(“身”)。
因此,丙酮酸通过有氧呼吸进入TCA循环代谢产能,可说是多细胞生物向愈发复杂化进化发展的必然选择。

说了这么多,TCA循环到底是怎么样的“圈圈”?它又是如何运转的?别急,这就来啦~~
作为胞质中糖酵解的最终产物,丙酮酸有以下几种去向:在细胞质中:可通过无氧呼吸生成乳酸(或乙醇)、或与谷氨酸在谷丙转氨酶(Glutamate pyruvic transaminase,GPT)催化下生成丙氨酸和α-酮戊二酸。在线粒体中:在能量充足的情况下,丙酮酸在丙酮酸羧化酶(Pyruvate carboxylase,PC)作用下生成草酰乙酸(Oxaloacetic acid,OAA),参与糖异生作用;在能量匮乏的情况下,丙酮酸则会优先转化成乙酰CoA,参与TCA循环,而这,就是我们故事的开端啦!
TCA的前期准备—丙酮酸变身乙酰CoA
华丽变身(丙酮酸脱羧加高能键--变成乙酰CoA)
丙酮酸在有氧状态下通过扩散作用穿过线粒体外膜,进入间隙中,再与位于线粒体内膜上的线粒体丙酮酸转运载体(Mitochondrial pyruvate carrier,MPC)结合,借助线粒体膜间隙的质子梯度(质子动力势)以协同运输的方式和质子一起转运到线粒体基质中,进而被基质内的丙酮酸脱氢酶系(pyruvate dehydrogenase complex,PDH)氧化成乙酰CoA。
等等,PDH是个啥?都包含哪些酶类? 这些酶类又是如何协同地介导丙酮酸脱羧呢?快一起往下看吧!
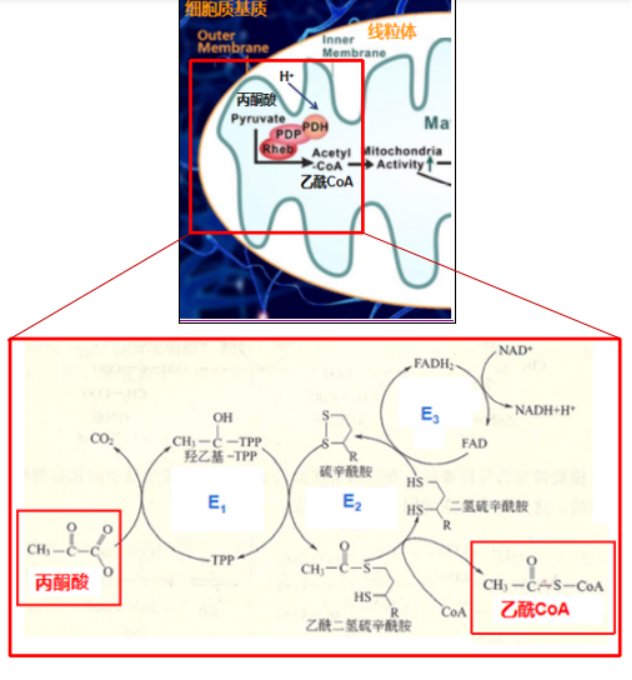
丙酮酸转换成乙酰CoA
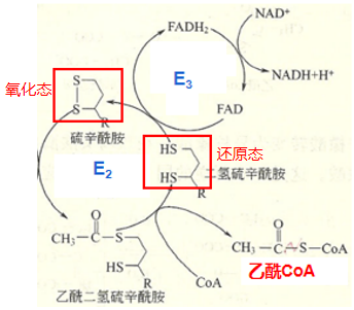
1.PDH构成:丙酮酸脱氢酶复合体(Pyruvate dehydrogenase complex,PDH)是多种酶和辅助因子通过非共价键结合在一起形成的多酶复合体,包含三种酶类:丙酮酸脱氢酶(Pyruvicacid dehydrogenase,PDH,即E1)、二氢硫辛酸转乙酰酶(Dihydrolipoyl transacetylase,DLAT,即E2)、二氢硫辛酸脱氢酶(Dihydrolipoyl dehydrogenase,DLD,即E3)以及六种辅助因子:焦磷酸硫胺素(Thiamine pyrophosphate,TPP)、硫辛酰胺(Lipoamide)、辅酶A (Coenzyme A,CoA)、黄素腺嘌呤二核苷酸( Flavin adenine dinucleotide,FAD)、烟酰胺腺嘌呤二核苷酸(Nicotinamide adenine dinucleotide,NAD+)以及镁离子(Mg2+)。
2.三步反应:丙酮酸转变为乙酰CoA的反应可以概括为以下三个步骤:丙酮酸的脱羧;碳单位的转移和氧化;氧化型硫辛酰胺的再生。反应将三碳(3C)的丙酮酸转变为二碳(2C)分子乙酰CoA。
咱们一步一步来看┗|`O′|┛ 嗷~~
第一步:丙酮酸的脱羧
E1借助TPP和Mg2+将丙酮酸(3C)的羧基脱去,生成CO2, 随后TPP接受反应后的乙酰基,乙酰基的羰基与TPP结合,发生加成反应,进而乙酰基变为羟乙基,生成了中间物羟乙基-TPP(2C)。
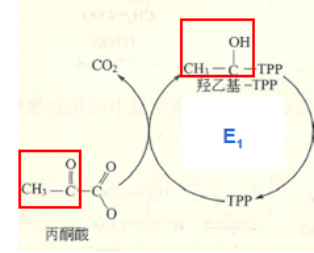
丙酮酸的脱羧示意图
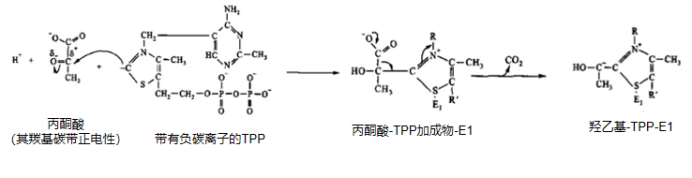
丙酮酸的脱羧反应式
第二步:碳单位的转移和氧化
在E2的催化下,硫辛酰胺接受羟乙基-TPP中的羟乙基,形成中间物(乙酰二氢硫辛酰胺),进而在E2的作用下,CoA接受乙酰基,转变为乙酰CoA,此时硫辛酰胺的氧化型二硫键(-S-S-)被还原成两个分开的巯基(-SH)。
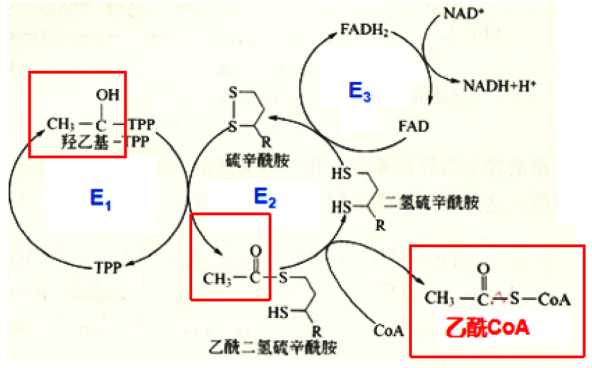
碳单位的转移和氧化示意图
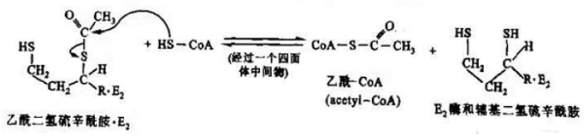
碳单位的转移和氧化反应式
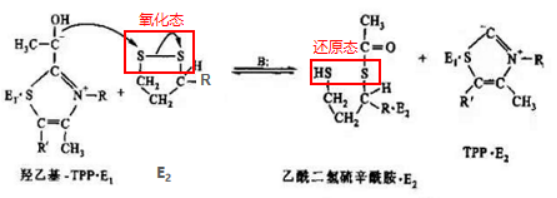
第三步:氧化型硫辛酰胺再生
随后,E3借助辅因子FAD,将还原型硫辛酰胺氧化成原来的形式,二硫键(-S-S-)再次形成,而FAD则被还原成了FADH2。之后,为了保证PDH反应前后功能上的一致,在E3的作用下将一分子的NAD+还原成NADH,使得FADH2又恢复成原本的氧化态FAD,继续在PDH中发挥氧化硫辛酰胺的功能。
氧化型硫辛酰胺再生示意图
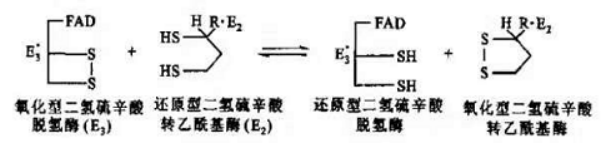
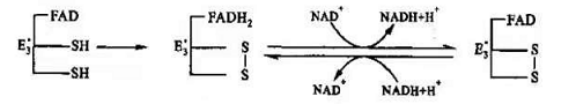
氧化型硫辛酰胺再生反应式
3. 调控:以上是丙酮酸进入TCA产能的前过程,生物能量利用的经济性使然,该反应受到严格而多样的调控,包括:PDH产物的竞争性反馈抑制(乙酰CoA和NADH);E1的共价修饰调节,例如,E1受到丙酮酸脱氢酶激酶(Pyruvate dehydrogenase kinase, PDK)和丙酮酸脱氢酶磷酸酶(Pyruvate dehydrogenase phosphatase,PDP)的磷酸化和去磷酸化修饰,磷酸化的E1为失活状态;而PDKs和PDP又受到别构调节,如细胞能量状态指示剂ATP、乙酰CoA和NADH能够激活PDKs,进而间接地抑制PDH的活性;而ADP和丙酮酸则能够抑制PDKs,并间接激活PDH的活性;胰岛素能够激活PDP,从而解除磷酸基团对于PDH的抑制作用。因此,PDH主要受到双酶的共价修饰调节,从而能够精确地根据细胞能量状态来影响糖代谢。

有PDH把关,TCA稳稳的
看到这里,你是否疑惑,PDH系统构成复杂,丙酮酸氧化脱羧步骤繁琐,细胞为什么不能简化反应?为什么不能让丙酮酸直接进入TCA循环中呢?

拓展阅读
PDH系统——TCA的闸门
1.不可逆反应保证了TCA的底物优先可得性。在高等动植物中,由PDH催化的丙酮酸生成乙酰CoA的过程是不可逆的,只有在某些能够利用CO2的微生物中,该过程才会可逆(见下文番外篇:“反向TCA”)。在有氧的状态下,丙酮酸在线粒体基质中会优先结合PDH,在PDH的催化下生成乙酰CoA。因此,由PDH所进行的将丙酮酸变为乙酰CoA的转换是糖有氧氧化中关键的不可逆反应,这一过程保证了TCA可以优先获得反应底物,为机体的代谢反应供能,在各个组织中均不可或缺。
另外,当细胞能量充足时(有丰富的碳骨架--线粒体基质内有过多的乙酰CoA时),乙酰CoA会反过来抑制PDH活性(产物的竞争性反馈抑制),并别构激活线粒体基质中的丙酮酸羧化酶(Pyruvate carboxylase,PC),在PC的作用下,丙酮酸转化成OAA,参与糖异生作用。肝脏是进行糖异生作用的主要器官,丙酮酸在肝细胞中既可以参与TCA,也能作为糖异生的底物,因此PDH利用丙酮酸的不可逆氧化和乙酰辅酶A对PDH和PC的调控,则利于肝细胞根据机体能量状态进行TCA并控制肝糖输出,在肝脏中是重要的反应。
2.多酶多反应步骤使得TCA的调控更精细。催化这个反应的PDH受到很多因素的影响,如反应产物乙酰CoA和NADH可以分别抑制酶系中的E2和E3的活性;如能量指示剂,ATP(表明能量充足)能够抑制E1的活性,而ADP(表明能量不足,需要生成ATP)和胰岛素(表明葡萄糖充足,可用来氧化产能)能够激活其活性。
PDH作为TCA的看门者,需要机体的严格把关。这一设计使得机体能够更精细的根据自身的营养需要判断分解代谢是否应该进行,从而有效地调节生物分子分解和合成代谢的平衡。

经过了PDH的洗礼,丙酮酸披上了华丽的外衣,转变成了高能分子乙酰CoA。乙酰CoA作为反应性强的高能小分子,不仅能够通过丙酮酸氧化脱羧产生;还能通过其他能源物质(如脂肪酸)分解代谢所产生,所以乙酰CoA可作为糖脂途径共同的代谢小分子参与TCA循环。
第一幕:脱羧阶段——CO2的释放
TCA循环中包含几个主要的中间代谢物是含有三个羧基的有机酸(柠檬酸、异柠檬酸和草酰琥珀酸),故被称为三羧酸循环(Tricarboxylic acid cycle,TCA cycle)),简称TCA循环。
TCA循环共包括8步反应,均在线粒体基质中进行。其中有三步不可逆,保证了TCA单向且有效的运转。最初,乙酰CoA作为一个二碳分子,加入到TCA的碳原子循环中,最终,以两分子CO2的形式被完全的代谢掉。过程所释放的自由能推进了TCA后续反应中碳单位的转移和重塑,产生更多的高能分子ATP(由氧化磷酸化生成)和GTP(由底物水平磷酸化生成)等。
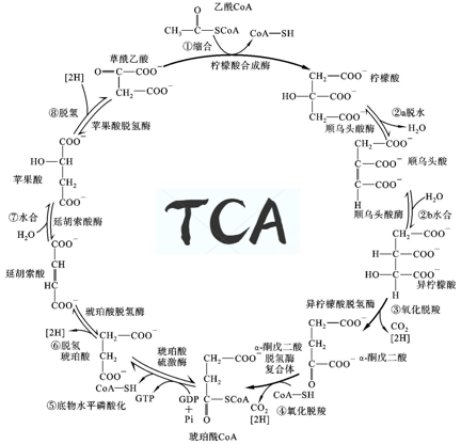
随着TCA循环中碳原子的转移,各个代谢分子都得到了更新和再生,这使得TCA循环和其他代谢途径的底物分配周转自如。
集中精力,
这是一场让人眼花缭乱的碳移位,满分操作来啦~

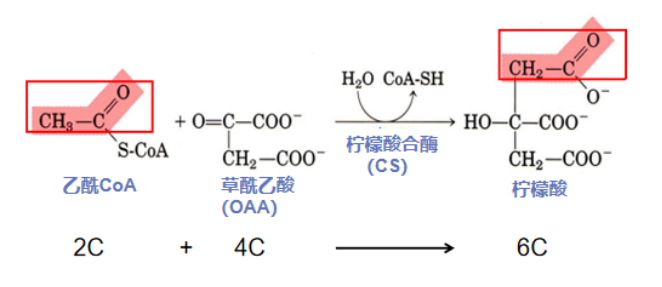
第一步:限速把关碳乘三(乙酰CoA与草酰乙酸缩合形成柠檬酸)
初入职场的乙酰CoA虽然只是一个含有高能键的“小小”二碳分子,却有着不小的野心和抱负,但踏入大门才发现自己一个大佬(酶)都不认识。草酰乙酸(Oxaloacetic acid,OAA)作为糖代谢的大红人,游荡在线粒体与细胞质基质中。它看到了乙酰CoA作为合资人的潜质,介绍给柠檬酸合酶(Citrate-synthase,CS)认识,乙酰CoA(2C)在伯乐OAA(4C)的游说下终于和CS握了手,最终和OAA合体,形成柠檬酸(6C)。

新人乙酰CoA
反应酶:柠檬酸合酶(Citrate-synthase,CS)是由两个亚基构成的二聚体,每个亚基的两个结构域构成一条较深的裂缝,其中含有草酰乙酸(Oxaloacetic acid,OAA)的结合位点。CS作为TCA循环第一步反应的催化酶,受到严格调控,该反应是不可逆的限速步骤。
反应流程:OAA与CS活性中心结合,使酶发生相应构象变化,从而创造出乙酰CoA的结合位点,CS的组氨酸残基掠取乙酰CoA的氢原子,使其碳原子不稳定,帮助乙酰CoA的负碳离子亲核进攻OAA羰基,二者偶联形成中间物(柠檬酰CoA),CS构象再次发生变化,天冬氨酸残基(Asp)切断柠檬酰CoA硫酯键,释放产物(柠檬酸和CoA)。反应释放的CoA可继续作为酰基载体,在氧化还原反应中被利用,而柠檬酸继续参与氧化分解。
拓展阅读
灭鼠药——氟乙酸
大家一定听说过毒鼠强,这是一种氟乙酸类鼠药,包括氟乙酰胺、氟乙酸和氟乙酸钠等。其灭鼠效果源于氟代乙酸是乙酸的类似物,能够被乙酰-CoA合成酶识别,在其催化下生成氟酰-CoA,氟酰-CoA能够与OAA缩合生成氟代柠檬酸而非柠檬酸。顺乌头酸酶(ACO)含有铁硫聚簇(Fe-S),而氟酰-CoA能够和Fe-S紧密结合,虽不是和酶活性中心共价相连,但结合力极强,氟酰-CoA“霸占”ACO活性中心,作为不可逆抑制剂抑制其活性,从而抑制TCA,造成全身能量耗竭,引起多脏器损害,达到灭鼠的目的,但由于死鼠中残存的氟乙酸仍会被其他动物接触,废水中也会残存,会造成环境污染和二次毒性,因此这种熟知的灭鼠药现已被禁用。
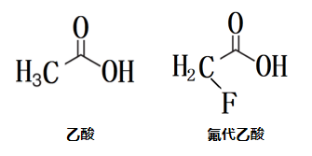
结构式对比
第二步:羟基异位谋发展(柠檬酸异构化形成异柠檬酸)
自从和OAA合体以后,乙酰CoA变大了,也变强了,大家都叫TA:“柠檬酸”。
柠檬酸在TCA部门混了个脸熟,却没有进一步发展。请教了一圈,其他分子都说:“要想融入TCA,就要保持周转,不能停滞不前。”于是职场小白柠檬酸决定改变自己,适应这里!柠檬酸本身不易氧化。因此,柠檬酸找到了造型师ACO帮忙打理外(结)表(构),一番脱水加水、羟基移位操作,成功变身为易于脱氢氧化的异柠檬酸。

我要变形啦
反应酶:乌头酸酶(aconitase,ACO)本质上是一个铁硫蛋白。含有由共价键结合的4个铁原子、4个无机硫化物和4个半胱氨酸硫原子形成的Fe-S聚簇。Fe-S聚簇可参与底物的脱水和再水合作用。
反应流程:柠檬酸的羟基处于第三位碳原子上,属于叔醇化合物,其本身的结构不利于氧化。为了参与下一步的氧化脱羧,柠檬酸在ACO作用下,先脱水再加水,使柠檬酸羟基由β碳原子转移到α碳原子上,实现了羟基的异位转变,羟基加在第二位碳原子上,成为了拥有伯醇基的异柠檬酸,为进一步的氧化脱羧反应作准备。
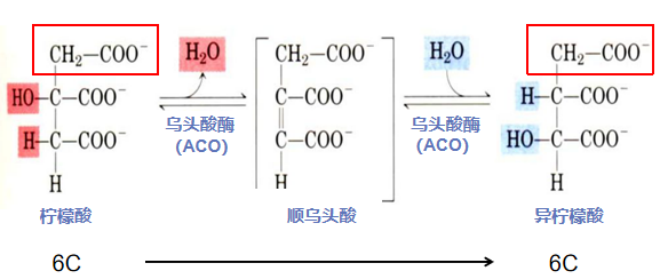
柠檬酸异构化形成异柠檬酸
拓展阅读
碳原子的系统命名法
在有机化学中,α-碳原子是用来标记碳原子顺序的,α位代表靠近官能团的第一位碳原子,以此类推,就依次命名为β,γ,Δ碳原子等等。若化学分子包含有多个官能团,则要考虑主链官能团的优先性去命名。一般主链官能团级别最高的是羧酸,其次是胺,再其次是醛酮,之后是醇,醚,炔,烯,烷等,卤素和硝基一般不考虑作为主链官能团。该命名方式有利于在众多复杂的化学结构中分析特定碳原子的性质,方便交流讨论。
反应的活泼程度与碳原子所处位置相关,α碳上的氢原子是极为活泼的,极易发生各种反应,其次为β碳上的氢原子,依此类推。因此,在TCA循环的第二步反应中,柠檬酸的氢原子位于β碳原子上,不易发生氧化作用,故借助ACO,将氢原子移位到α碳原子上,生成异柠檬酸,异柠檬酸α位的H原子较活泼,更易于进行第三步的氧化脱氢反应。
第三步:氧化脱羧裂-C-C-(异柠檬酸氧化形成α-酮戊二酸)
变身异柠檬酸后,机遇果然很快来了。异柠檬酸被IDH一眼看中,和他一起跑业务,并他的指点下和能量大佬NADH签成大单,为细胞供应大量紧俏的NADH。手握合同,回想着争取业务的辛劳过往,异柠檬酸(6C)长舒一口气,吐出CO2,变成了更轻盈更强大的α-酮戊二酸(5C)。
反应酶:异柠檬酸脱氢酶(isocitrate dehydrogenase,IDH)是多亚基变构调节酶,介导TCA连续两步氧化还原反应,先脱氢后脱羧。在真核生物线粒体中,IDH能够利用NAD+作为氢的受体,使得β-羟酸(异柠檬酸)氧化成β-酮酸(草酰琥珀酸),草酰琥珀酸结构不稳定,促进相邻C-C键的断裂,生成α-酮戊二酸,同时释放一分子CO2和一分子NADH。
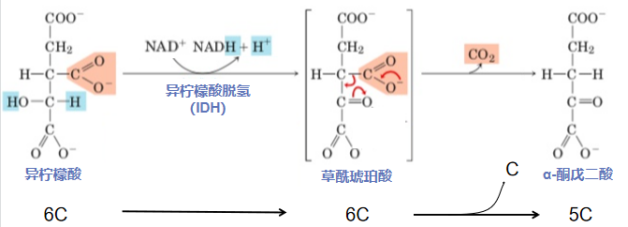
异柠檬酸氧化形成α-酮戊二酸
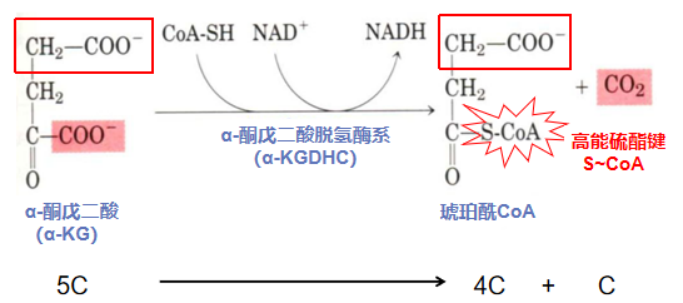
第四步:加能氧化再脱羧(α-酮戊二酸氧化脱羧形成琥珀酰CoA)
初战告捷后,α-酮戊二酸转到了大佬α-KGDHC手下干活。新工作很有挑战性,一开始进展不大,有同事开始在背后说闲话,说上次的NADH大单只是运气好。成熟的职场人当然不能受别人负能量的影响,深吸一口正能量(CoA-SH)的α-酮戊二酸(5C)再接再厉,又签到了NADH大单,扬眉吐气CO2,蜕变成拥有高能硫酯键的资深职场人琥珀酰CoA(4C)。

资深大佬气场就是不一样
反应酶:α-酮戊二酸脱氢酶系( α-ketoglutarate dehydrogenase complex,α-KGDHC)由三个酶 (α-酮戊二酸脱羧酶、硫辛酸琥珀酰基转移酶、二氢硫辛酸脱氢酶)和五个辅酶 (TPP、硫辛酸、CoA、NAD+、FAD)组成。该酶是催化TCA中第二次氧化脱羧反应,反应不可逆,需要NAD+和CoA作为辅助因子。
反应流程:在α-KGDHC催化下,CoA的高能硫酯基团转移氢原子到NAD+,NAD+形成NADH。CoA中不稳定的高能电子进攻羰基碳原子,导致-C-C-键断裂,生成带有高能硫酯键的琥珀酰CoA,释放1分子CO2,产生1分子NADH。

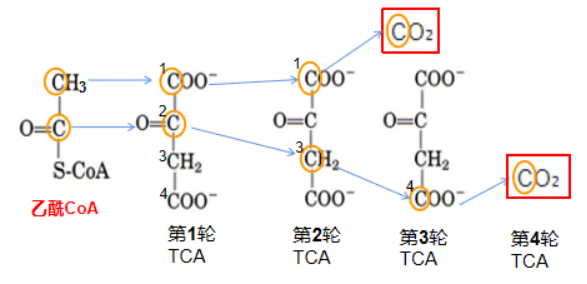
到这里为止,六个碳的柠檬酸共脱去两个羧基,释放出2分子CO2。聪明的你可能早已发现经过刚才脱羧阶段的四步反应,释放出的两分子二氧化碳中的碳原子都是来源于OAA,而乙酰CoA的碳原子仍完整保留(快倒回去看看,红框框里的属于乙酰CoA的两个碳原子是不是一直都没变?)。
实际上,TCA循环的八步反应可被大致分为两个阶段:脱羧阶段和再生阶段,到刚才的第四步为止,这轮TCA循环的脱羧反应已经全部结束了(即第三、四步反应),在这之后就再没有碳原子的丢失了。那么,最初来自乙酰CoA的两个碳原子要经历多少轮TCA循环才能完全代谢掉(即来自最初的乙酰CoA的两个碳原子全部以CO2的形式被丢失,不再存在于碳骨架体系中)呢?

答案:4轮
每一轮TCA循环均会生成两分子的CO2,第一轮TCA循环所释放的两分子C均来自于OAA(C1和C4),而乙酰CoA的两个C原子在第一次TCA循环的时候转变成了第二轮TCA中OAA的C1和C2;在进行第二轮TCA循环的时候,OAA的C1生成了CO2(来自乙酰CoA),C2变成了C3位;第三轮TCA循环时,OAA中的(来自乙酰CoA)C3位进一步变成C4位。在第四次TCA循环时,OAA中C4位的碳原子最终变成CO2释放出去,即完成了最初的乙酰CoA的碳原子的完全释放。因此乙酰CoA的两个C原子要经历4次TCA循环才能释放出去,从而被完全的氧化分解。
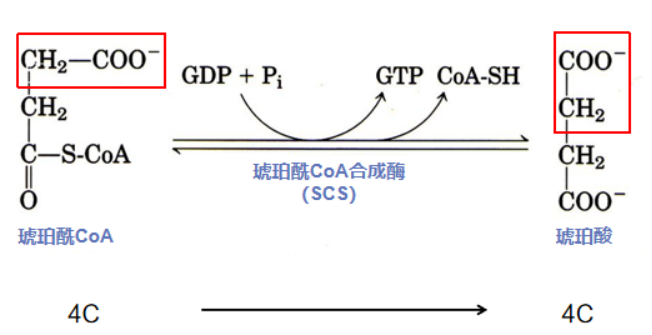
第五步:高能转移磷酸化(琥珀酰CoA转化成琥珀酸)
从职场小白(乙酰CoA)晋升为TCA高管的琥珀酰CoA,一入职便成绩斐然,利用自身的高能硫酯键与能量供应商GDP进行资源整合,产生了大批能量分子GTP,为TCA部门带来很大收益,琥珀酰CoA也因此再次晋升,成为TCA老大琥铂酸。
反应酶:琥珀酰CoA合成酶(Succinyl-CoA synthetase,SCS)由α和β两种亚基构成(分别由SucD和SucC两个基因所编码),在线粒体基质中,SCS通常以αβ二聚体发挥功能,Mg2+对酶的活性很重要。SCS催化琥珀酰CoA高能硫酯键水解是TCA循环中唯一一步底物水平磷酸化反应。
反应流程:在SCS的催化下,琥珀酰CoA拥有的高能硫酯键被水解,高能键断裂,释放的大量自由能通过底物水平磷酸化以GTP的形式储存下来,并转变成琥珀酸。反应生成的GTP可直接利用,也可以和ATP相互转换,此外,GTP还可以提供鸟嘌呤核苷酸用于DNA复制和转录反应。
小总结:从上述TCA的五步反应中我们可以看出,这个过程中释放了2分子的CO2,两分子NAD+被还原成NADH,同时还通过底物磷酸化产生了一分子高能磷酸基团(GTP)。哦,对了,十分重要的一点是,该过程产生的NADH后续又可以通过线粒体内膜上的电子传递链被氧化成NAD+,一方面完成NAD+的循环再生,另一方面参与线粒体膜间隙质子梯度的建立,为合成更多ATP分子做出贡献。
回顾了下目前的成果,可谓收获颇丰。其他乙酰CoA听说自己的同辈在TCA部门混得很好,也纷纷前来报道。但是,之前协助乙酰CoA融入部门的伯乐OAA已经蜕变为琥珀酸了。面对后续涌入二碳分子们,TCA部门又该如何保证TA们能够被持续氧化代谢,使得能量持续产生呢?

抢OAA啦!
这里就要 cue TCA循环里的“循环”二字啦!循环是以乙酰CoA和草酰乙酸的缩合生成柠檬酸为起始。现在乙酰CoA源源不断的进入,打配合的草酰乙酸不够了,怎么办?不用担心,小小的细胞极具智慧,接下来的重头戏就是琥珀酸如何重返初心、再生为草酰乙酸,达成循环之旅啦!生命体通过形成循环实现底物和酶的再利用,既体现了经济性,又能够实现对能量代谢的整体调控,瑞思拜!
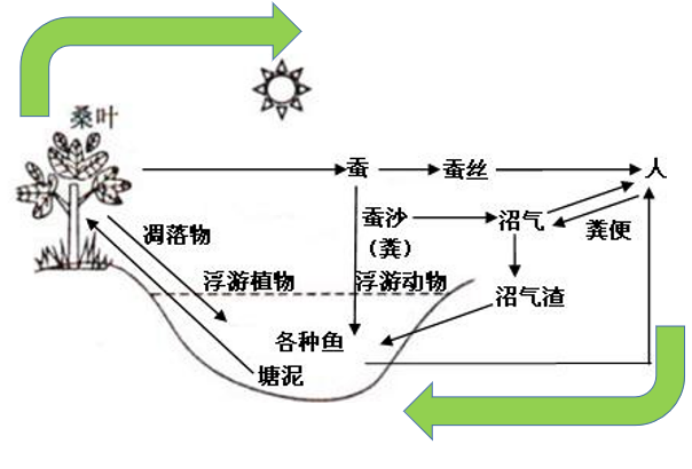
第二幕:达成循环——OAA再生

TCA循环转起来!
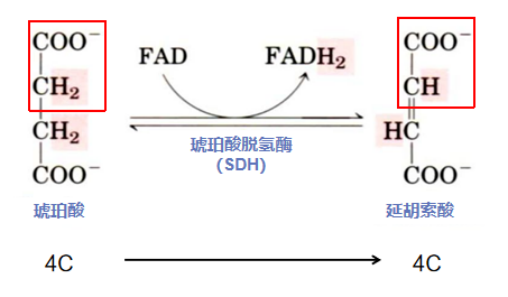
第六步:脱氢放能在膜上(琥珀酸脱氢形成延胡索酸)
反应酶:琥珀酸脱氢酶(Succinate dehydrogenase, SDH)属于氧化还原酶,是 TCA循环中唯一一个整合在线粒体内膜上的多亚基酶。FAD+作为SDH的辅基,其第8位碳原子与酶的组氨酸残基通过共价键的形式连接。
反应流程: SDH以FAD+作为琥珀酸脱去氢离子的受体,介导琥珀酸两个碳原子之间的氧化,即碳-碳键(-C-C-)的氧化脱氢。
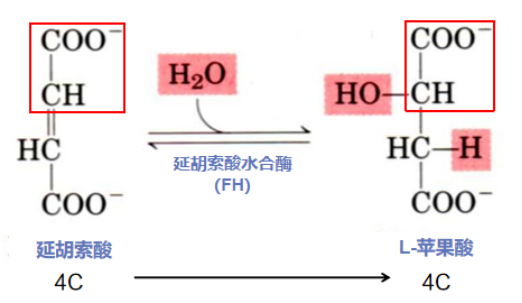
第七步:加成水合获羟基(延胡索酸水合形成L-苹果酸)
反应酶:延胡索酸水合酶(Fumarase hydratase,FH)的催化反应具有严格的立体专一性,只能催化一种构型的底物。该反应过程的-OH严格地加在靠近延胡索酸双键的一侧,而另一个-H加在相反的另一侧,从而只能形成单一构象的产物。由于FH活性中心只能和一种异构体匹配,结合/生成单一结构的底物/产物(L-苹果酸),使得FH相对于其他酶类显得比较特别,TCA循环中的其他酶类只区分底物类型,对于底物的异构体并无要求。
反应流程:在一分子H2O参与下,延胡索酸酶的活性中心特异性地结合底物延胡索酸,催化延胡索酸双键打开,发生加成反应,生成L-苹果酸,其具有β-碳原子位置上的羟基,从而为OAA的再生做准备。
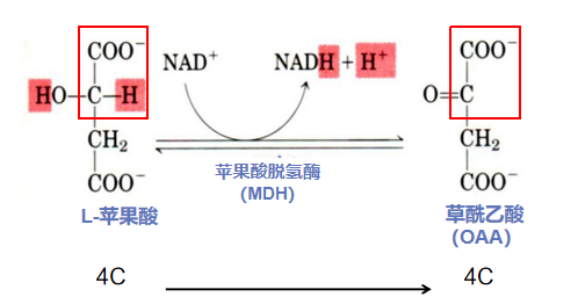
第八步:脱氢放能重归位(L-苹果酸脱氢形成草酰乙酸)
反应酶:苹果酸脱氢酶(malate dehydrogenase,MDH)作为TCA循环中最后一个氧化还原酶,辅酶为NAD+。作为唯一一步可逆的氧化反应,该过程的正反应标准自由能变化ΔG>0,在热力学上是不利于进行的,但由于TCA第一步反应中:OAA和乙酰CoA反应自由能变化ΔG<0,是高度放能的反应,非常利于进行,因此OAA作为第一步的底物更容易被消耗,使得第八步的正向反应虽然不易进行,但随着产物OAA的消耗(乙酰CoA与OAA的结合,记不清的小伙伴请倒回去看第一步~),正反应也得以进行,即一个热力学上不利的反应(苹果酸生成OAA)借助与其相偶联的热力学上的有利反应(乙酰CoA与OAA结合的放能反应)的推动而变得可行。
反应流程:MDH氧化L-苹果酸,使其羟基形成羰基,转变成OAA,NAD+作为氢原子受体,生成NADH。此步骤达成了OAA的再生和TCA循环的闭合。重新生成的OAA可以和乙酰CoA再次结合形成柠檬酸,参与新一轮的TCA循环。
哦,对了,细胞能量充足时,OAA还可以跨过线粒体内膜,进入到细胞质基质参与糖异生,具体将在第三幕介绍哦~
L-苹果酸脱氢形成草酰乙酸
小结
以上就是三羧酸循环的全部反应,让我们来分析一下:
1、四次氧化脱氢,生成三分子NADH、一分子FADH2;
2、两次脱羧,生成两分子CO2;
3、一次底物水平磷酸化,产生1分子GTP;
4、8步TCA反应中存在三种限速酶(CS、IDH、α-KGDHC),介导三步不可逆的限速反应。
整个TCA循环的8步反应包括4次氧化脱氢、2次脱羧、1次底物水平磷酸化、1次异化、水合、缩合反应。由TCA介导的4步氧化脱氢反应发生在线粒体内部(分别是第3,4,6,8步,其中第3,4,8步发生在线粒体基质;第6步在线粒体内膜上),其产生的NADH和FADH2可以直接进入位于线粒体内膜的氧化呼吸链进行电子传递。根据对呼吸链中质子的贡献程度,NADH可以再产生2.5个ATP分子;FADH2可以再产生1.5个ATP分子。

一分子乙酰CoA参与TCA循环,过程中产生3分子NADH、1分子FADH2、1分子GTP。
根据对电子呼吸链中质子的贡献程度,NADH可以产生2.5个ATP分子;FADH2可以产生1.5个ATP分子。
因此:3 x NADH:3 x 2.5ATP
1 x FADH2:1 x 1.5ATP
1 x GTP:1 x 1ATP
总计:10 分子ATP
计算2:一分子丙酮酸经TCA循环代谢后,共产生多少ATP分子?
根据以上结果,一轮TCA循环产生10 分子ATP。再加上丙酮酸转变成乙酰CoA的前期准备过程(生成1分子NADH=2.5ATP),即一分子丙酮酸参与一轮TCA循环共产生12.5分子ATP。
总计:12.5 分子ATP
计算3:一分子葡萄糖经TCA循环代谢后,共产生多少ATP分子?
一分子葡萄糖首先要经历10步糖酵解反应(在细胞质基质中),净生成2分子NADH、2分子ATP和2分子丙酮酸。
其中,细胞质基质中的NADH需要借助位于线粒体内膜的穿梭途径才能进入线粒体电子传递链,甘油-3-磷酸穿梭途径(主要存在于脑和骨骼肌)将NADH转变成FADH2输送到线粒体基质;苹果酸-天冬氨酸穿梭途径(主要存在于肝和心肌)则可以直接将NADH输送到线粒体基质。因此,胞质中的1分子NADH经历甘油-3-磷酸穿梭途径产生1.5个ATP分子,或经历苹果酸-天冬氨酸穿梭途径产生2.5个ATP分子。

棕色脂肪中存在甘油-3-磷酸穿梭途径,甘油-3-磷酸穿梭途径转换能量较低,多余能量可用于棕色脂肪产热过程。
试想一下,若葡萄糖不参与TCA,直接通过糖酵解转化成不完全氧化产物的话,只会产生2分子ATP。小小葡萄糖蕴含这么多的能量,我们如果不通过TCA来物尽其用,充分发挥它的价值,那我们岂不是亏大了!

中场休息
由于篇幅限制,《三羧酸循环之丙酮酸升职记》到这里先中场休息一下,等道具师换幕布和道具,我们下周同一时间再见,敬请关注噢~
关注微信公众号代谢学人
了解更多前沿资讯

https://m.sciencenet.cn/blog-3483272-1314877.html
上一篇:代谢学人--Cell & Nature 近期代谢研究精选
下一篇:代谢学人--三羧酸循环之华山论剑