博文
代谢学人——Nature Metabolism:“PE”脂代谢让肌肉健康起来
||
代谢学人
Nature Metabolism:"PE"脂代谢让肌肉健康起来
撰文 | 闪光余 马莹 张俊 仲银召 郭明伟
编辑 | 孟美瑶
校对 | 张俊

背景介绍
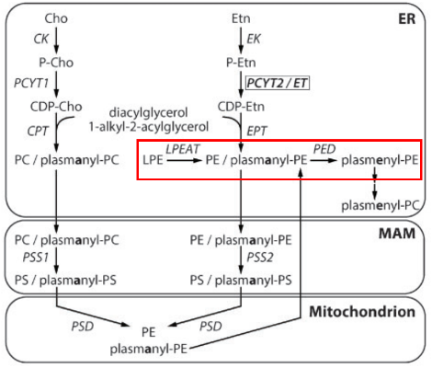
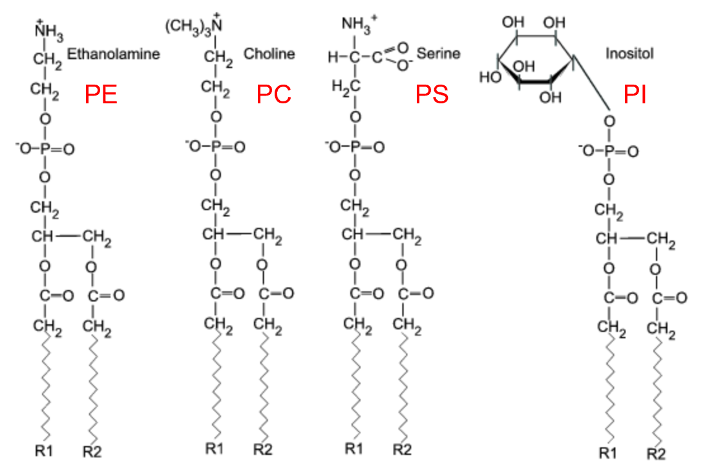
骨骼肌是人体最大的器官之一,在机体的支撑、运动和代谢方面起着重要作用。遗传性疾病、慢性疾病或衰老导致的肌肉退化严重损害了数百万人的生活质量和健康,因此,弄清肌萎缩的形成机制是现代医学的一个重要目标。细胞中真核脂质组是极为复杂的,可产生多达10万种不同的脂质,并且特定脂质之间的差异体现在亚细胞区室、细胞和组织类型的分布上。具体而言,例如,甘油磷脂的生物合成上存在亚细胞区室差异,如磷脂酰乙醇胺(Phosphatidylethanolamine,PE)和磷脂酰胆碱(Phosphatidylcholine,PC)在内质网中合成,而磷脂酰丝氨酸(Phosphatidylserine,PS)则在线粒体中合成。此外,在组织分布上,PE缩醛磷脂(PE etherphospholipids,PE[O],PE的甘油主链的sn-1位置含有乙烯醚连接的烷基链)在大脑中含量最丰富,它对大脑的灰质代谢和白质结构具有正向作用,保护神经元膜和髓鞘免受氧化损伤的危害。总之,组织特异性差异表明某些器官使用特定的脂质途径来促进器官健康和长寿。
正常的脂肪代谢对肌肉健康十分重要。例如,此前有文献报道一种名为中性脂肪沉积症(neutral lipid storage disease,NLSD)是一种罕见的常染色体隐性遗传病,主要由脂肪代谢途径中的酶或辅基缺陷导致三酰甘油(triglyceride,TG)在不同组织细胞内异常沉积引起,可导致中性脂肪沉积症伴肌病(neutral lipid storage disease with myopathy,NLSDM),表现为肌无力、肌肉疼痛、运动后易疲劳等明显的肌肉症状。此外,在人类的一种脂质从头合成通路(Kennedy pathway)中,合成PE的关键酶胞苷转移酶PCYT2/ECT的遗传缺陷导致复杂且严重的遗传性痉挛性截瘫(HSP)。在机制上,由于PCYT2的遗传缺陷,细胞中缺乏正常水平的CDP-乙醇胺,所以无法将磷酸乙醇胺基团转移到二酰甘油的sn-2羟基上,抑制了PE或PE[O]的形成,进而导致大脑中进行性神经元丢失(progressive neuronal loss),极大抑制了中枢神经系统的发育和功能。尽管PE的合成还存在两条补偿途径,即PS脱羧为PE和外源补充Lyso-PE后再酰化为PE,但是临床研究表明,这些补偿途径并不能完全挽救PCYT2遗传缺陷造成的HSP表型。
在本篇文章中,作者发现了PCYT2合成的PE在肌肉健康中的特定作用。pcyt2突变斑马鱼和肌肉特异性Pcyt2敲除小鼠表现出生长停滞,身材矮小,肌肉发育受损和进行性虚弱,加速衰老和缩短寿命。相比之下,在其他组织中敲除Pcyt2的小鼠似乎不受影响。肌肉中PCYT2的敲除导致线粒体和细胞脂质组的改变,影响线粒体功能和脂质双分子层的物理化学性质,进一步损害肌膜稳定性和运动耐力。作者在文中进一步表明,在衰老的人和小鼠肌肉中,PCYT2活性下降,并且针对Pcyt2的基因治疗可改善Pcyt2敲除和衰老小鼠的肌肉力量。因此,PCYT2和通过PCYT2合成的PE对于肌肉健康是必不可少的,并将线粒体和肌细胞膜的脂质与肌肉退化、运动耐力和衰老联系起来。

敲黑板啦!
1. 缺乏PCYT2的个体表现出生长停滞;
2. PCYT2的缺陷会导致线粒体功能丧失;
3. PCYT2的缺陷会导致肌肉生理生化功能稳定性的降低;
4. 肌肉特异性治疗PCYT2缺陷小鼠会改善肌肉衰老。
研究结果
1 具有致病性PCYT2突变的个体生长停滞
最近在具有复杂疾病的个体中发现了PCYT2突变,这些复杂疾病包括发育性运动迟缓和进行性整体肌肉无力。在这些突变个体中,作者发现在PCYT2中具有纯合无义突变(NM_001184917.2:c.1129 C > T,p.Arg377Ter)的那些个体从出生起,在整个童年期和成年期表现出明显较低的体重和较短的体长(图1 a,b)。作者还评估了具有SELENOI/EPT1(编码Kennedy途径中合成PE的最终酶)突变的个体(扩展图1),这些个体表现出PCYT2突变相似的临床特征。事实上,在EPT1中具有纯合变体(NM_033505.4:c.335 G > C,p.Arg112Pro)的个体也表现出生长缺陷,进一步证实了Kennedy途径在出生后生长中的作用。因此,除了先前描述的症状之外,在Kennedy途径中产生PE的两种关键酶的突变与发育不良有关。
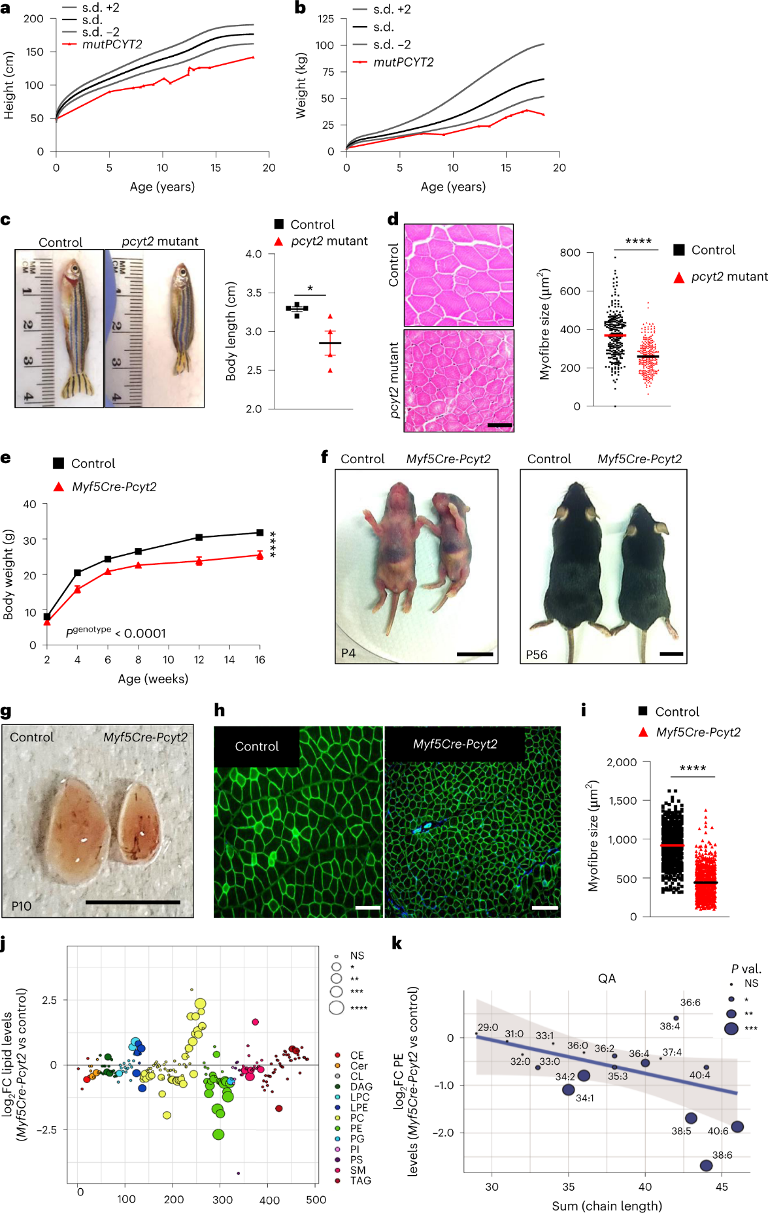
图1 人类PCYT2罕见疾病突变和pcyt2突变斑马鱼的表型
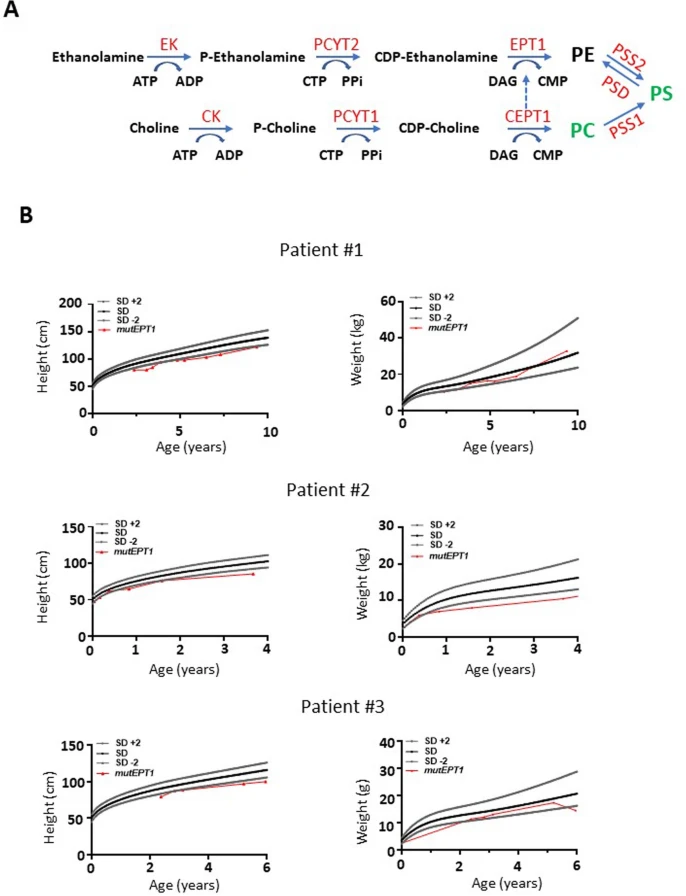
扩展图1 PE合成途径与EPT1罕见病突变个体常规饮
2 斑马鱼pcyt2基因缺陷影响肌肉和全身生长
由于文献表明Ept1的缺失可以被Cept1(PC合成通路的最终酶,但Cept1可以发挥Ept1的功能,进而合成PE)部分代偿,而PCYT2在各个组织中普遍表达,负责催化PE形成,敲除该基因可能会导致复杂且严重的疾病,模拟人体的PCYT2突变,因此作者将研究重点放在了PCYT2上。为了了解病理机制,作者首先在斑马鱼上突变pcyt2 。类似在人类中出现的罕见疾病症状,pcyt2突变的斑马鱼体型明显较小(图1 c)。肌肉的发育对整个身体的生长至关重要,肌肉萎缩的一个显著特征是生长停滞。因此,作者检测了pcyt2突变斑马鱼的肌肉形态。他们发现其骨骼肌更小,肌纤维更小,但总数没有变化(图1d和扩展图2a,b)。这些结果表明,肌肉发育受损可以解释斑马鱼和人类中与pcyt2功能缺失突变相关的生长迟缓。
拓展阅读
Kennedy途径和甘油磷脂
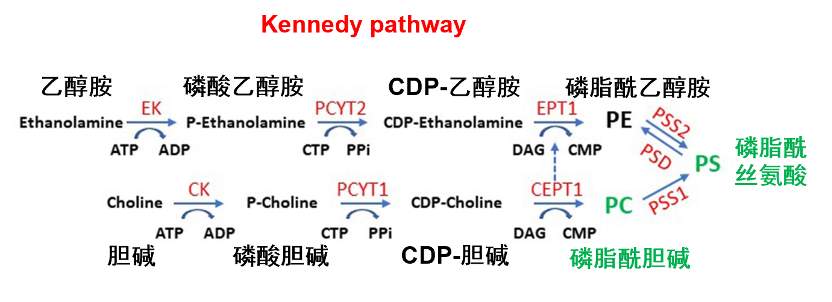
甘油磷脂构成了膜脂的基本成分,占据了整个膜脂的50%以上。甘油磷脂为3-磷酸甘油的衍生物,包括磷脂酰胆碱/卵磷脂(phosphatidylcholine,PC)、磷脂酰丝氨酸 (phosphatidylserine,PS)、磷脂酰乙醇胺(phosphatidylethanolamine,PE)和磷脂酰肌醇(phosphat-idylinositol,PI)等,主要在内质网合成。组成生物膜的甘油磷脂分子的主要特征是具有一个与磷酸基团相结合的极性头和两个非极性的尾(脂肪酸链),但存在于线粒体内膜和某些细菌质膜上的心磷脂除外,它具有 4个非极性的尾部。在分布上,PC位于磷脂双分子层外小叶,而PS、PE、PI则主要分布于磷脂双分子层内小叶。在功能上,四种甘油磷脂作为细胞膜的主要成分之一,可以保持细胞膜的完整性和稳定性,防止细胞膜的破裂和溶解。然而,由于其噒酸基团又被各种结构不同的小分子化合物酯化,因此它们的功能也存在一定差异。(1)PE能够与多种氨基酸残基建立氢键,进而抑制胞质侧的阴性氨基酸的定位,是膜蛋白跨膜结构域拓扑结构的重要决定因素。此外,PE还具有促进收缩环的拆卸(细胞分裂)和膜融合(细胞融合)的功能;(2)PC是肝细胞正常极低密度脂蛋白分泌所必需的,并且也是各种第二信使的主要来源,进而参与细胞信号转导;(3)PS可以通过与其他蛋白质相互作用来调节细胞信号传导,从而影响细胞的增殖和分化。此外,PS还可以参与凝血过程,调节血小板的聚集和血管的收缩。(4)PI可以通过调节细胞内钙离子的浓度来影响细胞的代谢和信号传导。
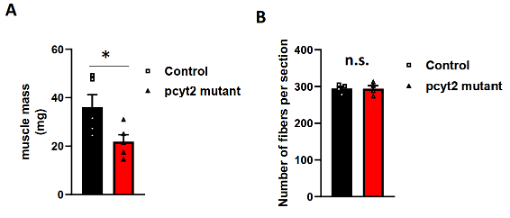
扩展图2 pcyt2突变斑马鱼的肌肉表型
3 小鼠中的Pcyt2肌肉缺陷损害肌肉和全身生长
在小鼠中,Pcyt2的全身敲除会导致胚胎致死。因此,为了研究Pcyt2在肌肉健康中的作用,作者构建了骨骼肌特异性Pcyt2敲除小鼠,即将Pcyt2fl/fl小鼠与Myf5启动子驱动的Cre小鼠杂交以产生Myf5-Cre-Pcyt2杂交后代,由于Myf5是骨骼肌形成的第一个广泛表达的成肌调节因子(MRFs),因此作者可以在小鼠的肌肉生长早期模拟人类HSP疾病。通过RNA测序验证了Pcyt2基因缺失(扩展图3a)。Myf5-Cre-Pcyt2小鼠以正常孟德尔比率出生,但与对照相比,在出生P1和生长早期P4时体形较小,体重增加较少,并且在出生后生长较慢(图1e,f和扩展图4a-d)。而Myf5-Cre小鼠和Pcyt2fl/fl小鼠对照均未出现以上表型,因此,作者使用Pcyt2fl/fl同窝小鼠作为所有后续实验的对照。此外,作者注意到Myf5-Cre-Pcyt2小鼠在P10和2月龄时的四肢肌肉较小(图1g和扩展图4e-h),骨骼肌纤维面积减小(图1g-i)。从10日龄Myf5-Cre-Pcyt2小鼠分离的QU的脂质组学分析显示,PE,特别是长链脂肪酸PE的水平显著降低(图1 j,k)。此外,研究人员发现Kennedy通路中磷脂酰胆碱(PC)分支来源的PC生成增加,并且Kennedy通路中的PE和PC分支的几种酶的表达上调(小编注:PC途径和PE途径中的酶在底物使用中存在一定重叠,由于KO小鼠中的PE途径缺乏一种关键酶,所以PE无法正常合成,底物的堆积和反馈机制激活了PE和PC途径其他酶的活力,同时使PC含量上升。但是二者功能不完全相同,已在前文拓展阅读详细介绍)(图1j和扩展数据图4i)。
拓展阅读
肌肉特异性Cre
Myf5-Cre:Myf5是与调节骨骼肌谱系分化高度相关的转录因子,在骨骼肌和卫星细胞表达,Myf5+细胞敲除后会出现严重的肌肉损失。小鼠肌源因子5(Myf5)启动子驱动骨骼肌谱系中干细胞、成肌细胞和肋骨中一些软骨祖细胞中靶基因和cre的表达;此外BAT细胞来自Myf5细胞谱系,因此Myf5-Cre通常用于研究棕色脂肪组织的发育和成肌过程。
HSA-cre:HSA为人源a-肌动蛋白启动子,人与小鼠同源,Cre活性仅限于体细胞和心脏的成人横纹肌纤维和胚胎横纹肌细胞。
MCK-cre:是肌酸激酶启动子,这种转基因小鼠具有由肌酸激酶启动子驱动的Cre重组酶基因,在心肌和骨骼肌中都高表达。
以上三种肌肉特异性的Cre重组酶都可以用于构建肌肉特异性敲除的小鼠模型。
参考文献:
[1] Zhao Q, et al.Proc Natl Acad Sci USA.2020 Sep8; 117(36).
肌肉生长过程中,肌肉卫星细胞增殖使肌纤维数量增多,P7之后发生肌纤维肥大。在Myf5-Cre-Pcyt2小鼠的肌肉和体外成肌细胞中,作者发现增殖细胞的数量和成肌细胞的增殖比例未受影响(扩展图5a,b)。尽管Pax7+肌肉干细胞的数量和分布比例在成年敲除小鼠和对照小鼠中相似(扩展图5c),但是敲除小鼠肌肉的成肌细胞融合显著减少,并且肌纤维更细小(图2a-c)。促融合基因和晚期分化基因的表达下降也验证了上述结果(图2d)。文献表明,膜脂质组成对于成肌细胞融合是必不可少的。鉴于PE脂质是丰富的膜脂质,于是作者想要探究去除成肌细胞膜上的PE是否会影响成肌纤维融合。作者使用无毒的PE特异性SA-Ro结合探针在成肌细胞融合期间占据外部暴露的PE(小编注:SA-Ro结合探针即链霉亲和素(SA)偶联的PE结合肽。正常状态下,PE分布在细胞膜脂双层内小叶上发挥功能,如在细胞分裂中促进收缩环的拆卸,而SA-Ro可特异性与PE结合形成一个复合物,破坏了脂双分子层的不对称性,进而抑制PE发挥正常功能),模拟PE缺乏。结果表明,成肌细胞融合受到严重影响(图2e-g),表明膜PE是成肌细胞融合所必需的(小编注:PE、PC和PS都属于磷脂,其中PE和PS位于脂双分子层的内小叶,PC位于外小叶)。
拓展阅读
肌纤维形成过程中的分化基因
肌细胞最初出现时是以单核的形态出现的,在后来逐渐进化成多核,多核肌细胞形态的出现为动物提供了强大、灵活的肌肉组织,而肌细胞从单核到多核形态的发生伴随着肌细胞融合的过程,在该过程中,基因的表达也发生着变化。成肌细胞融合主要涉及3个关键步骤:首先成肌细胞彼此识别并粘附在一起,其次细胞膜的脂质双层发生重排形成融合孔,最后融合孔扩张形成合胞体并出现多核肌管。这三个步骤是受到高度调控的动态过程。
许多调控因此参与了细胞融合的过程,如MRFs(肌源性调节因子)家族,如:MyoD、Myf5、MyoG和Myf4。MRFs家族的两个成员MyoD和Myf5的表达由Pax基因激活,该基因还可以增强肌肉前体细胞的增殖。MyoD和Myf5是第一批在肌肉中表达的MRFs,触发肌细胞的分化,被认为是肌形成的决定因子;而MyoG和Myf4在分化后期高度表达,触发成肌细胞融合最终形成肌管。除了分化基因的表达之外,还有融合基因也调控成肌纤维融合的过程。Myomaker和Myomerger是两种必需的肌肉特异性融合蛋白(由MYMK和MYMX编码),它们共同在非融合细胞中重建融合,Myomaker参与脂质双层重排过程,其被认为能有效促进肌源性细胞融合,它还可以促进非融合细胞(如成纤维细胞和成肌细胞融合)间的融合,但并不能促进非肌细胞间发生融合。而Myomerger对于融合孔的形成是必要的。
参考文献:
[1] Deng S. Semin Cell Dev Biol. 2017 Dec;72:45-55.
[2] Lehka L. Semin Cell Dev Biol. 2020 Aug;104:81-92.
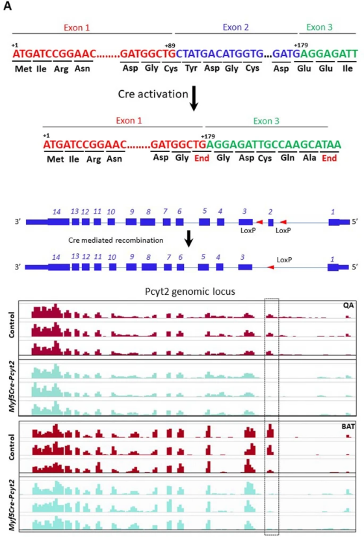
扩展图3 小鼠Pcyt2敲除分析
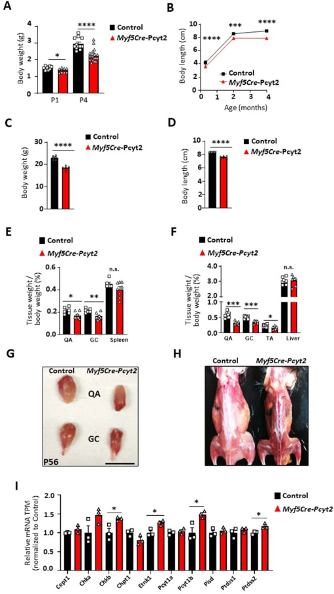
扩展图4 Myf5-Cre-Pcyt2小鼠的表征
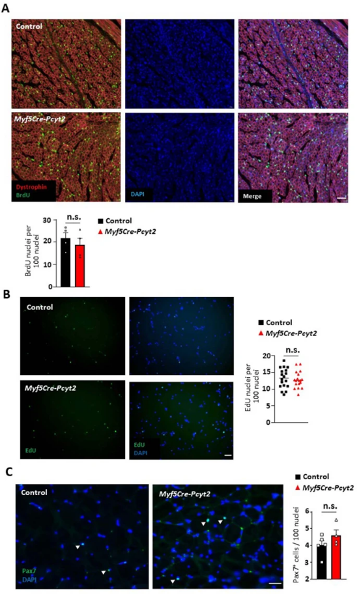
扩展图5 Myf5-Cre-Pcyt2小鼠成肌细胞增殖
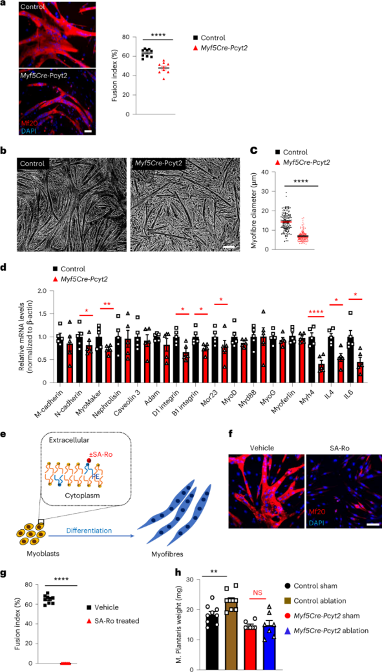
图2 Pcyt2缺陷影响肌肉干细胞融合和肌肉肥大性生长
4 缺乏Pcyt2的肌肉表现出进行性肌萎缩
作者注意到成年Myf5-Cre-Pcyt2小鼠表现出尾悬时后肢扣紧(hind limb clasping following tail suspension),表明存在肌无力(图3a,b)。此外,随着敲除小鼠的衰老,其肌肉力量逐渐下降(图3c和扩展图7a)。在敲除小鼠8月龄时均表现出驼背(图3d),同时该表型也出现在人类PCYT2相关疾病中(图3e)。并且Myf5-Cre-Pcyt2小鼠肌肉组织发生明显萎缩(图3f),细胞核中心化更高(图3g)。此外,作者观察到12至15月龄的敲除小鼠肌肉中的管状聚集体(小编注:密集排列的膜性包涵体区域,来源于肌浆网和线粒体膜,其内含有具有多种酶活性的各种蛋白质,肌肉出现管状聚集体则会导致疼痛、痉挛或僵硬)和炎症(图3h和扩展数据图7b-d)。由于观察到的肌肉无力在两种性别中都很明显,Myf5-Cre-Pcyt2小鼠出现继发性骨质减少(图3i),导致整体虚弱和寿命缩短(图3j)。
拓展阅读
肌再生细胞核中心化
肌纤维是成熟骨骼肌的基本单位,其中细胞核位于每个肌纤维的外围。但是在一些患有肌病的患者肌肉中表现出大量中心化细胞核的肌纤维,这些肌病称为中央核肌病(CNM)。近50年来,中心化细胞核一直被用作区分肌肉疾病和神经系统疾病的病理标志物,此外,细胞核中心化也是肌纤维修复的标志,在此期间,新生肌纤维细胞核都经历了至少三个剧烈的核运动、包括集中、对齐、扩散、外围迁移和锚定。与稳态肌肉相比,当肌肉进行修复时,更多的肌纤维将具有中心位置的细胞核。因此,中心位置的细胞核提供了一种简单的测定方法,以确定哪些肌纤维正在因疾病或肌损伤而进行修复。
参考文献:
[1] Folker ES, Baylies MK. Front Physiol. 2013 Dec 12;4:363.
此外,进行性肌萎缩对全身代谢都有显著影响。在6月龄时,葡萄糖清除率随着血糖水平和食物摄入量的逐渐下降而增加(扩展图7e-f、8a,b),这可能是死亡率增加的部分原因。在室温下,尽管Myf5-Cre-Pcyt2小鼠不活跃(扩展图7g),但是其光照期间和黑暗期间的能量消耗明显增加(扩展图7h)。在热中性状态下,敲除小鼠在光照期间的能量消耗与对照小鼠相当,而在暗期间,能量消耗更大(小编注:本文敲除小鼠肌肉萎缩,进食量减少,但整体能量消耗是升高的,作者对此也没有给出解释,作为排除因素之一,在下一部分探究了敲除小鼠的BAT产热能力没有明显变化)(扩展图7i)。并且,敲除小鼠的活动量降低和肌无力也十分明显(扩展图7j,k)。
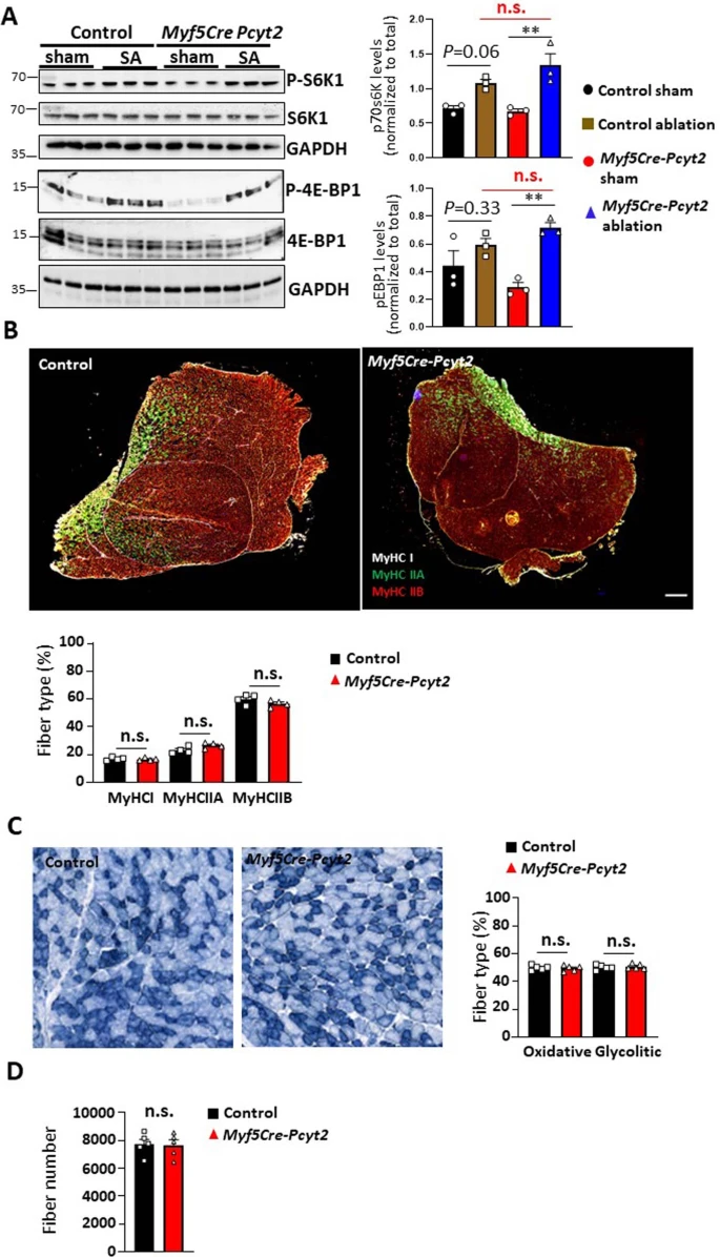
扩展图6 Myf5-Cre-Pcyt2小鼠骨骼肌肌纤维类型分布
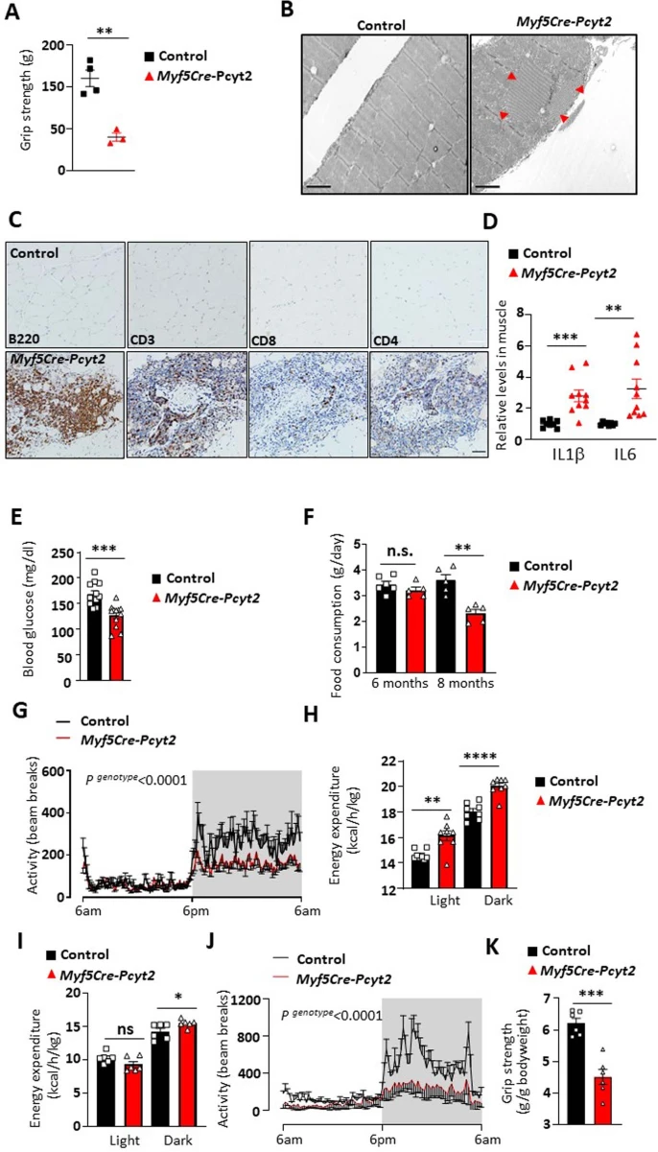
扩展图7 Myf5-Cre-Pcyt2小鼠的肌肉炎症和代谢评估
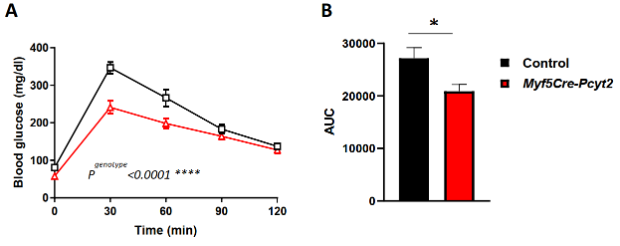
扩展图8 6月龄Myf5-Cre-Pcyt2小鼠葡萄糖耐量实验
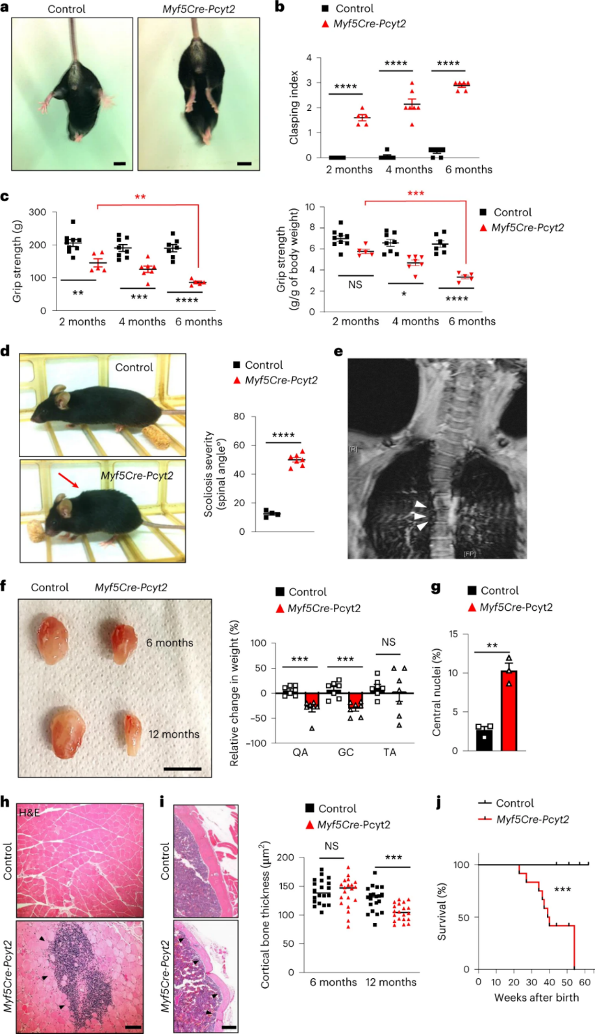
图3 小鼠中Pcyt2的失活导致进行性虚弱、肌肉萎缩、炎症和加速衰老
5 Pcyt2在骨骼肌中至关重要
Myf5同时在骨骼肌和棕色脂肪组织(BAT)的发育过程中高表达。作者发现PE脂质在Myf5-Cre-Pcyt2小鼠的BAT中减少,但程度显著低于骨骼肌,同时PC脂质增加(扩展图9a,b)(小编注:今年2月一篇Sci Adv报道了PE在BAT线粒体的UCP1依赖性产热中发挥重要作用,PC则报道较少)。重要的是,Pcyt2的缺失并不影响脂肪前体细胞体外分化为棕色脂肪,不影响通过BAT活性的体温调节,也不影响BAT中Ucp1的水平(扩展图9c-f)。并且在Myf5-Cre-Pcyt2小鼠的BAT中,线粒体超微结构、含量和功能基本正常(扩展图9g-j)。此外,作者将Pcyt2fl/fl小鼠与AdipoQ-Cre小鼠杂交来特异性敲除白色和棕色脂肪组织中的Pcyt2(小编注:Myf5-cre在前体细胞阶段敲除目标基因,AdipoQ-cre则在成熟脂肪细胞中敲除目标基因),但是并没有观察到生长、血糖相关或任何明显病理学表型(扩展图10a-c)。除此之外,在运动神经元(Mnx1-Cre)、肠上皮(Villinl-Cre)、乳腺和皮肤上皮细胞(K14-Cre)中敲除Pcyt2,在12个月龄时都没有明显的发育或退行性影响(扩展图10d-l)。以上结果表明Pcyt2在这些组织中不起重要的发育作用。
为了进一步探索肌肉特异性敲除Pcyt2的影响,作者将Pcyt2fl/fl与Mck-Cre小鼠杂交。与作者在Myf5-Cre-Pcyt2小鼠和人类PCYT2突变个体中的发现的表型相反,已报道Mck-Cre-Pcyt2小鼠在年轻时具有有益效果。关键的膜肌发育在生命早期就已完成,通过肌肉卫星细胞和成肌细胞的增殖和融合来促进。但是Mck-Cre在肌肉形成的后期活性最高,同时肌肉成熟后并不会存在剧烈的成肌细胞增殖和融合(小编注:运动一般会促进肌肉细胞增殖和融合,但该品系小鼠没有做运动模型相关实验),加之膜PE周转缓慢,共同解释了观察到的相反表型。重要的是,尽管4个月龄的年轻Mck-Cre-Pcyt2小鼠不显示退行性表型,但是衰老的MckCre-Pcyt2小鼠显示出明显的肌无力(扩展图10m-n)。
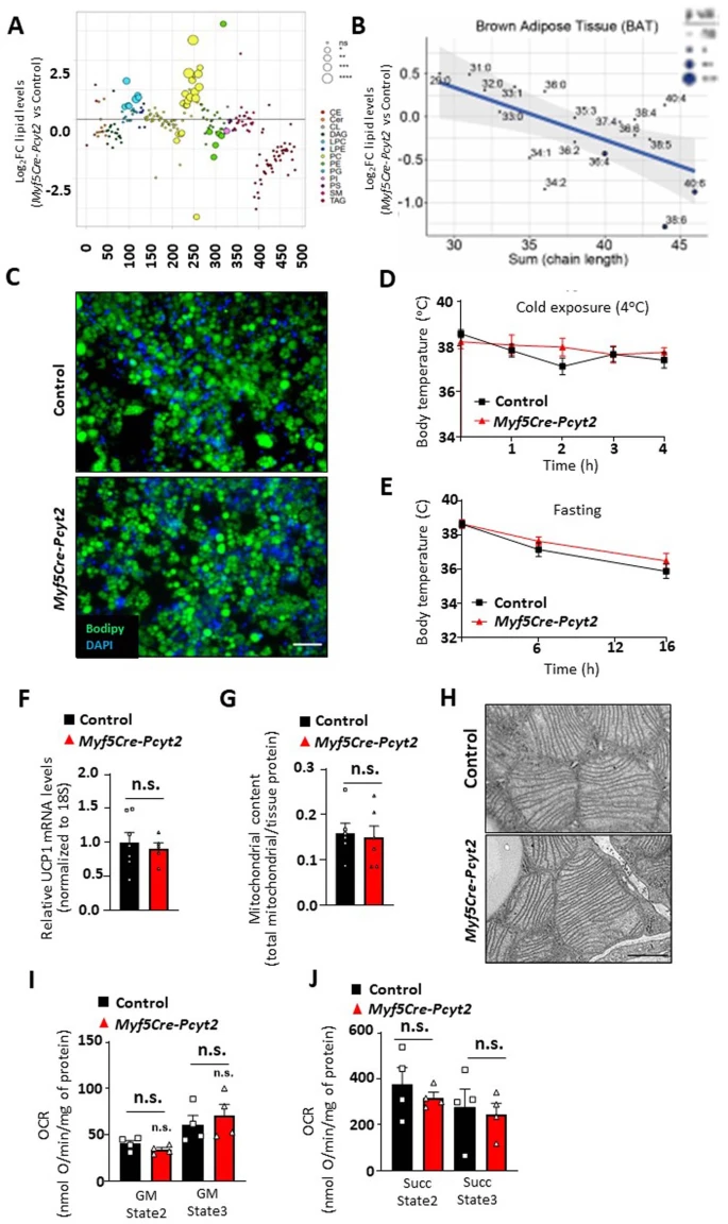
扩展图9 Myf5-Cre-Pcyt2小鼠棕色脂肪组织的表征
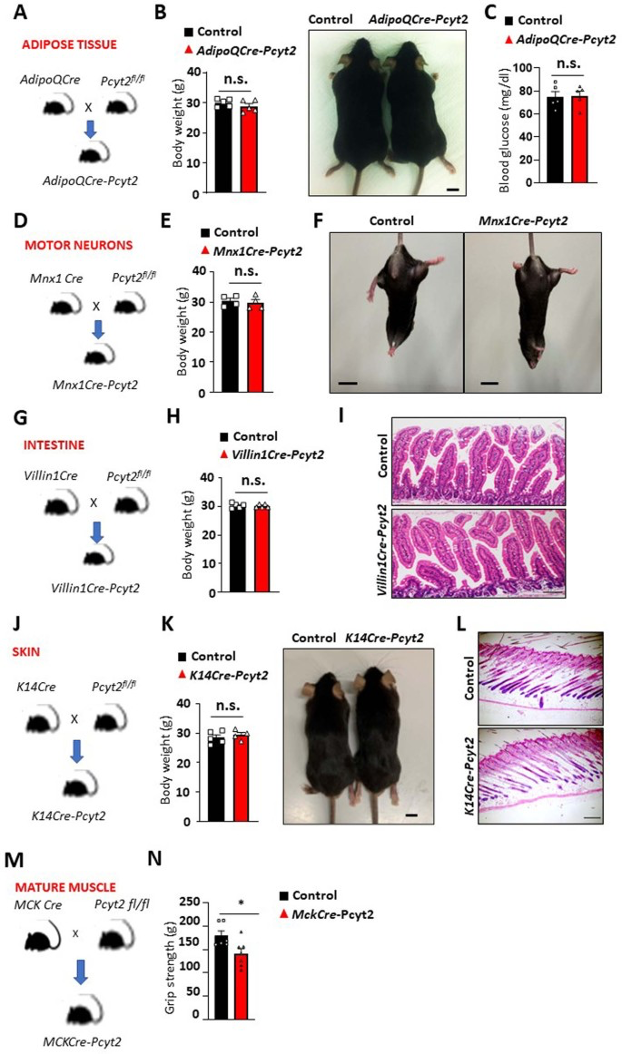
扩展图10 Pcyt2在多种小鼠组织中的特异性失活
6 Pcyt2肌肉敲除改变线粒体功能
鉴于PE在细胞膜和线粒体膜中丰富,作者研究了Pcyt2缺乏是否影响线粒体稳态。与全组织脂质组变化相似,来自Myf5-Cre-Pcyt2小鼠的肌肉线粒体中也存在PE脂质的减少(图4a,b)。Myf5-Cre-Pcyt2小鼠的肌肉RNA转录组分析显示与线粒体功能障碍相关的基因得到富集(图4c)。有趣的是,当作者分析来自2月龄Myf5-Cre-Pcyt2小鼠的骨骼肌线粒体活性时,作者观察到活性增加,随后来自6月龄Myf5-Cre-Pcyt2小鼠的活性下降(图4d和扩展图11 a-c)。此外,作者观察到分离的肌纤维中线粒体活性氧(mtROS)的增加,以及6月龄Myf5-Cre-Pcyt2小鼠的骨骼肌中抗氧化剂过氧化氢酶活性和蛋白质氧化损伤水平的增加(图4e-g)。同时,细胞应激的标志物(pJNK,Foxo1)和肌萎缩的标志物(Atrogin,MuRF1,Fbx031)在6月龄敲除小鼠的肌肉中增加(图4h,i)。即使在具有明显表型的成年突变小鼠中,线粒体的超微结构形态和含量在肌肉中也没有变化(扩展图11d,e)。因此,增加的细胞应激包括能量应激、ROS介导的蛋白质损伤和不平衡的蛋白质稳态加速Myf5-Cre-Pcyt2小鼠的肌肉变性(小编注:作者认为 2月龄小鼠的线粒体可能被过度激活,因此表现出活性即耗氧量增加,进而引发ROS诱导的损伤,并在6月龄表现出肌萎缩)。
接下来,作者测试了Pcyt2底物磷酸乙醇胺的积累是否影响线粒体。与对照线粒体相比,磷酸乙醇胺对从Myf5-Cre-Pcyt2小鼠分离的骨骼肌线粒体的活性没有抑制作用(扩展图11f),表明线粒体中过度积累磷酸乙醇胺不是骨骼肌线粒体活性降低的原因。接着,作者试图通过从4月龄开始每天施用线粒体靶向抗氧化剂SS-31,持续2个月来改善Myf5-Cre-Pcyt2小鼠的病理特征。尽管握力和组织重量有轻微改善,但效果不显著(扩展图11g,h)。
作者还测试了Pcyt2和Pcyt2催化形成的PE在肌膜Ca2+摄取/释放和自噬中的潜在作用。作者未能观察到6个月龄敲除小鼠分离肌纤维中Ca2+释放和Ca2+摄取的任何结构变化(扩展图12a-c),这表明肌浆网Ca2+处理得到保留。文献表明,PE与ATG8的偶联对于自噬也是必需的。然而,在进食和禁食条件下,小鼠四头肌和膈肌中的PE-ATG8偶联体无明显变化,并没有明显的p62/SQSTM1积累(小编注:p62是被广泛研究的自噬底物。在自噬体形成过程中,p62作为链接LC3和聚泛素化蛋白之间的桥梁,被选择性地包裹进自噬体,之后被自噬溶酶体中的蛋白水解酶降解,所以p62蛋白的累积表征自噬活性的下降和自噬体异常堆积)(扩展图12d-i),表明敲除Pcyt2对肌细胞自噬无明显影响。值得注意的是,Myf5+祖细胞(可双向分化为肌肉细胞和棕色脂肪细胞)中敲除Atg7或抑制LC3的PE脂化会诱导棕色脂肪过度活化,但不导致肌肉无力、变性或肌肉营养不良,与作者的Myf5-Cre-Pcyt2小鼠相反。总之,Pcyt2的敲除不影响自噬或肌浆网中的PE功能,但影响线粒体功能。然而,由于线粒体靶向疗法没有挽救Myf5-Cre-Pcyt2小鼠的肌肉无力,这表明存在另外的细胞缺陷导致前文的病理学特征。
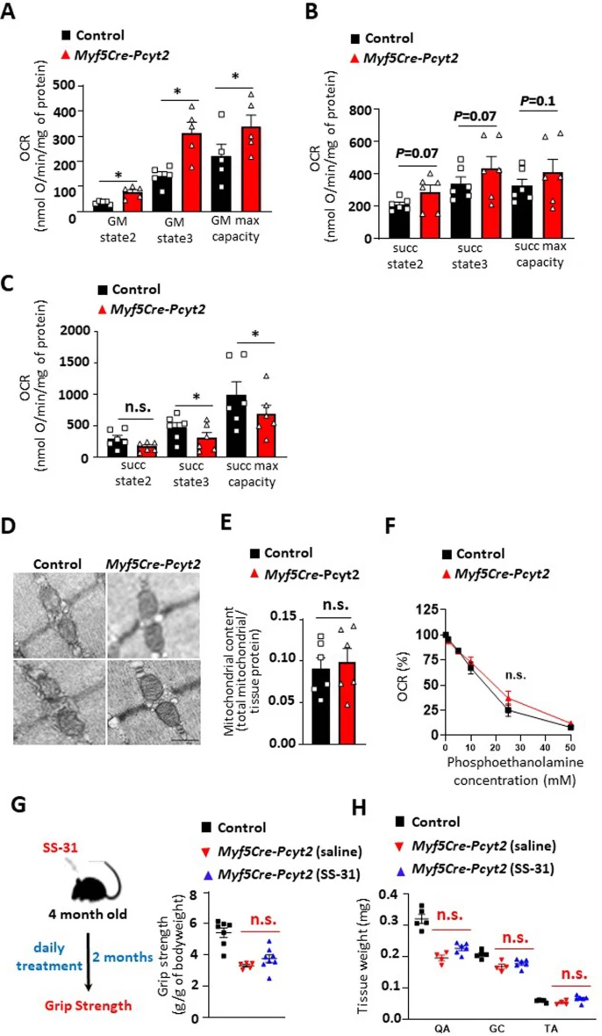
扩展图11 线粒体稳态评估和SS-31治疗
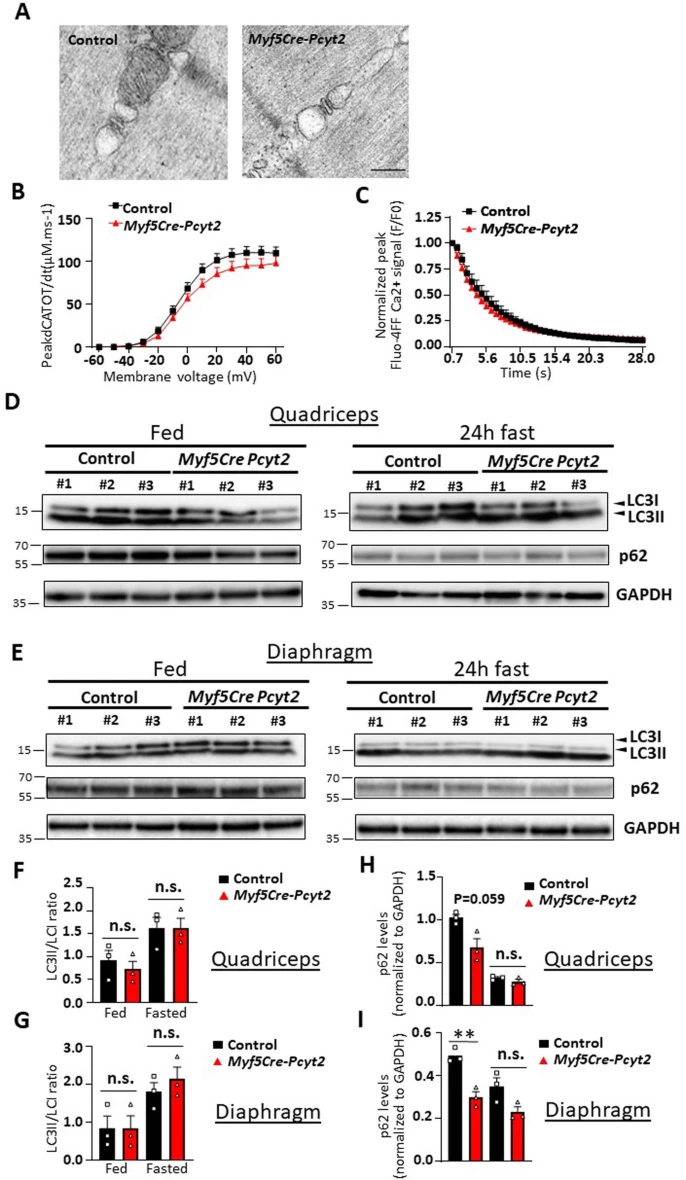
扩展图12 骨骼肌中钙处理和自噬标志物的评估
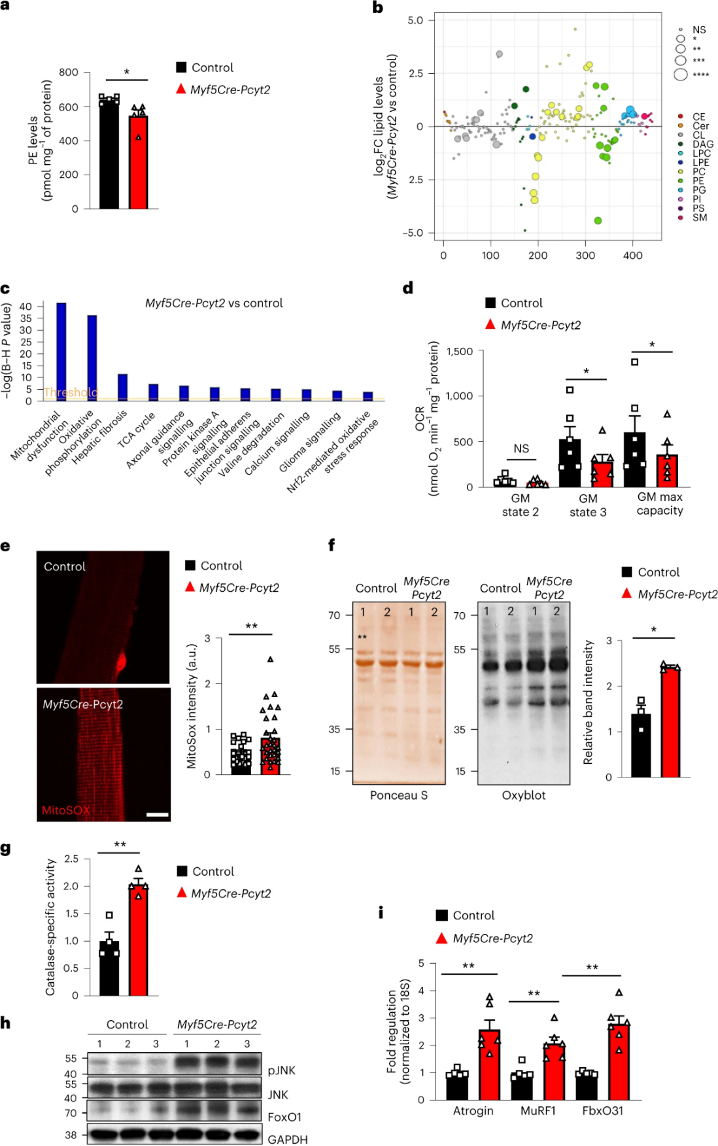
图4 与棕色脂肪线粒体相反,Pcyt2缺乏严重影响肌肉线粒体稳态
7 Pcyt2缺陷影响肌膜脂双层理化性质
肌膜稳定性受损导致肌营养不良症中的肌纤维变性。作者的肌肉全组织脂质组学数据显示含有长链FA的PE减少,它是丰富的膜脂质。因此,作者假设Myf5-Cre-Pcyt2小鼠中含有长链FA的PE丰度降低也可能影响肌膜脂质双层的形成和稳定性,从而驱动肌肉病理学。为了验证这一假设,作者首先评估了Myf5-Cre-Pcyt2小鼠的肌膜脂质双层的组织是否改变。NR12S染色的巨质膜囊泡(GPMV)的光谱成像提供了脂质双层在其天然组成复杂性和结构组织中的结构信息(小编注:为了研究和模拟细胞膜的结构和性质,可采用化学试剂诱导法使细胞分泌巨型细胞膜囊泡GPMV,来源于细胞的巨型囊泡尺寸与细胞相似,又具有细胞膜的结构特点,它作为一种天然的仿生材料在细胞膜研究以及构建仿生药物递送系统等研究领域具备独特的优势)。作者推导出一般极化(GP)的参数,较高的值对应于一个紧密包装、刚性的脂质双层,较低的值对应于一个松散包装的软双层。引人注目的是,与对照成肌细胞衍生的GMPV相比,衍生自Myf5-Cre-Pcyt2成肌细胞的GPMV显示松散堆积和较软的脂质双层(图5a,b)。为了进一步探究膜双层变化是否持续存在于肌纤维中,作者在从组织分离后立即评估了来自肌纤维的GPMV(图5c)。从Myf5-Cre-Pcyt2小鼠的肌纤维分离的GPMV再次显示脂质堆积的显著减少和软脂质双层(图5d)。
为了直接探究膜脂质双层的这些结构-化学变化如何影响机械性质,作者在分离的肌纤维上使用高分辨率布里渊光散射显微镜(BLSM)。使用BLSM的扫描显示,与对照相比,从Myf5-Cre-Pcyt2小鼠分离的肌纤维的表面硬度显著降低(图5e,f)。单个肌纤维上的原子力显微镜进一步证实,与表达Pcyt2的肌细胞相比,Myf5-Cre-Pcyt2肌纤维在施加纳米级水平的压力后具有更高程度的膜变形和更低的膜刚度(图5g,h)。因此,在成肌细胞和肌纤维中敲除Pcyt2导致膜脂质双层的结构改变,直接扰乱肌膜脂质双层的刚性和刚度的机械性质。

图5 Pcyt2缺失导致肌膜结构改变
8 Pcyt2缺陷损害肌膜稳定性
完整的细胞膜结构对膜屏障功能至关重要。骨骼肌肌膜在收缩过程中容易形成损伤,需要结构稳定以达到正常的骨骼肌功能。为了确定Myf5-Cre-Pcyt2小鼠中肌膜的扰动结构是否导致渗透性的改变,作者在6月龄对照和Myf5-Cre-Pcyt2小鼠中腹腔注射伊文思蓝。相对于对照,作者观察到伊文思蓝在Myf5-Cre-Pcyt2小鼠的四头肌中的广泛积累(图6a,b)(小编注:轻微渗透是正常存在的,但是损伤越严重渗透越厉害)。为了进一步探索肌膜稳定性,作者在新鲜分离的肌纤维上诱导激光介导的膜微损伤,并通过测量荧光FM1-43染料的细胞内流入来真实的定量损伤程度。在激光微损伤后,Myf5-Cre-Pcyt2肌纤维表现出渗透性增加(图6c)。
为了直接探究体内肌膜对肌损伤的影响,作者对Myf5-Cre-Pcyt2小鼠进行离心运动来诱导肌损伤。结果表明,敲除小鼠在前期适应阶段(4m/min,40min,持续3天)和中期训练阶段(4m/min,40min;9m/min,20min;持续7天)的运动表现与对照一致,然而,在后期应激阶段(20m/min,20 min,持续2天),敲除小鼠未能完成运动(图6d)(小编注:前期、中期强度不大,小鼠能够顺利跑完,但也形成损伤。KO小鼠修复能力较差,所以当后期强度提高后运动表现差于WT鼠)。此外,在对照小鼠中,训练后四头肌和腓肠肌的重量较高,而敲除小鼠的肌肉没有经历肥大过程(扩展图13a);相反,它们的骨骼肌表现出明显的肌肉损伤,并出现炎症、脂肪细胞沉积和纤维化(图6e-f,扩展图13b)。与此同时,作者观察到敲除小鼠运动后血液中肌酸激酶水平的增加(图6g),表明其损伤更严重。肌膜相关膜修复蛋白Dysferlin在训练后异常定位于Myf5-Cre-Pcyt2四头肌中(扩展图13c)(小编注:Dysferlin是一种细胞骨架蛋白,主要位于肌细胞膜下表面)。此外,作者在训练后观察到敲除小鼠中的F-肌动蛋白网紊乱(小编注:F-肌动蛋白细胞骨架组成成分,紊乱代表细胞形态异常,功能受损)(图6h)。值得注意的是,两种小鼠的骨骼肌中Pax7+肌肉干细胞的数量无明显差异(扩展图13d),表明敲除小鼠的损伤修复能力受损并非在于干细胞的增殖分化上。因此,Pcyt2的PE合成是肌膜稳定性所需的,从而促进离心运动后的肌损伤修复过程。
拓展阅读
离心运动
肌肉的收缩方式有多种,根据锻炼时肌肉形态不同,可分为向心收缩和离心收缩。骨骼肌在相同运动负荷下,离心收缩比向心收缩产生更大的力,同时离心运动由于存在高负荷和低能量需求的特点,故十分适合运动损伤后或不能进行传统抗阻力量训练的人群。
离心收缩,其特征是肌肉-肌腱复合体的延长,当施加在肌肉上的力超过肌肉本身产生的瞬时力时,就会产生肌肉离心收缩(也称为延长收缩),导致肌肉肌腱系统在收缩时被迫延长,在这个过程中,肌肉吸收由外部负荷产生的能量,因此离心作用被称为“负功”,吸收的机械能可以作为热量散失,或者能量可以回收并施加到随后向心作用期间产生的主动力中。在离心运动过程中,肌肉容易发生损伤和延迟性疼痛,称为迟发性肌肉酸痛(DOMS)。因此,离心运动可作为诱导肌损伤模型之一。
参考文献:
[1] Douglas J, Sports Med. 2017 Apr;47(4):663-675.
[2] Hody S, Front Physiol. 2019 May 3;10:536.
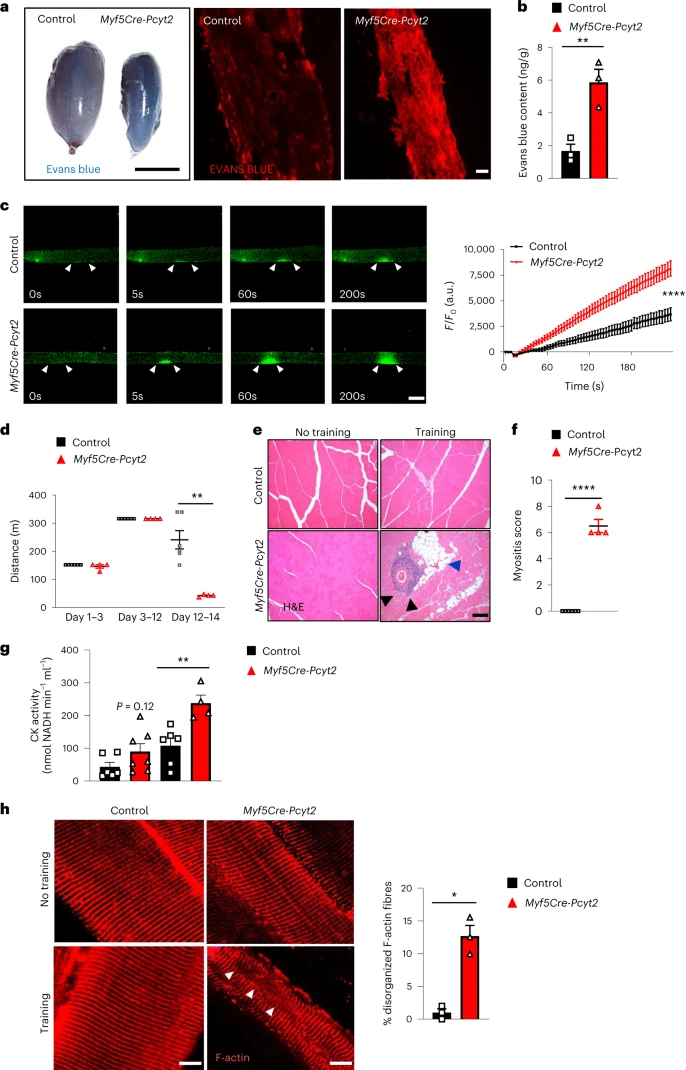
图6 Pcyt2对肌肉细胞膜的完整性和损伤修复至关重要
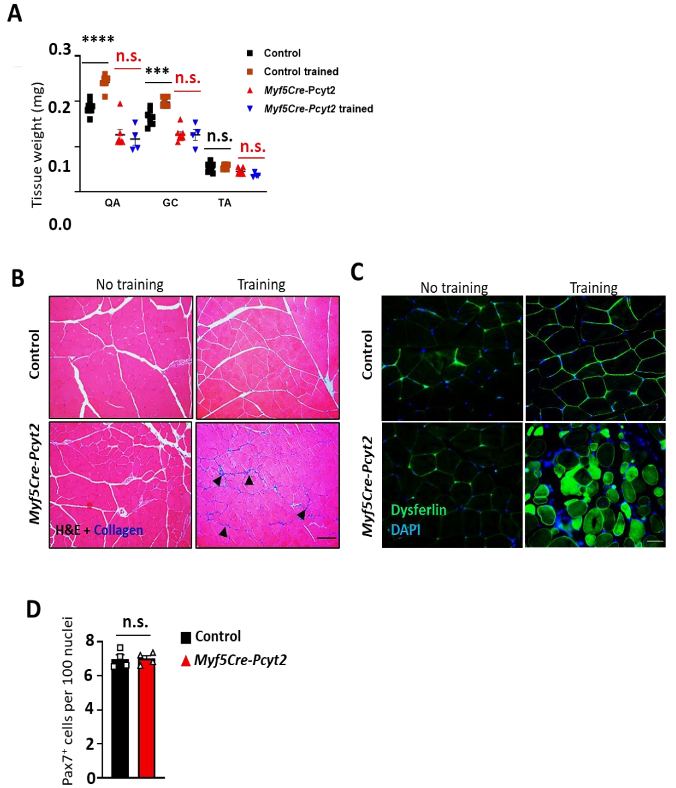
扩展图13 Myf5-Cre-Pcyt2小鼠偏心运动后肌肉质量、肌肉损伤和Pax7+干细胞数量
9 肌肉特异性Pcyt2基因治疗改善突变小鼠的肌无力
目前,没有针对PCYT2缺乏的疾病的疗法。由于基因疗法在治疗罕见疾病方面取得了显着进展,作者试图在疗法上进行创新,进而改善Myf5-Cre-Pcyt2小鼠的肌无力。作者将在肌肉肌酸激酶8(CK8)启动子/增强子的控制下的Pcyt2克隆到腺相关病毒(AAV)6载体中(小编注:CK8能够特异性靶向肌肉组织)。将AAV6-CK8-Pcyt 2病毒腹腔注射到4日龄的Myf5-Cre-Pcyt2小鼠中,并在6个月后评估这些小鼠的肌肉(图7a)。与未处理的Myf5-Cre-Pcyt2对照相比,该方法导致治疗组小鼠的骨骼肌中Pcyt2蛋白过表达和PE水平上升(图7b-c,扩展图14a)。此外,治疗组的突变小鼠握力改善、骨骼肌质量和肌纤维直径增加(图7d-f,扩展图14b),骨骼肌表现出改善的肌膜特征和线粒体功能(图7g-i)。
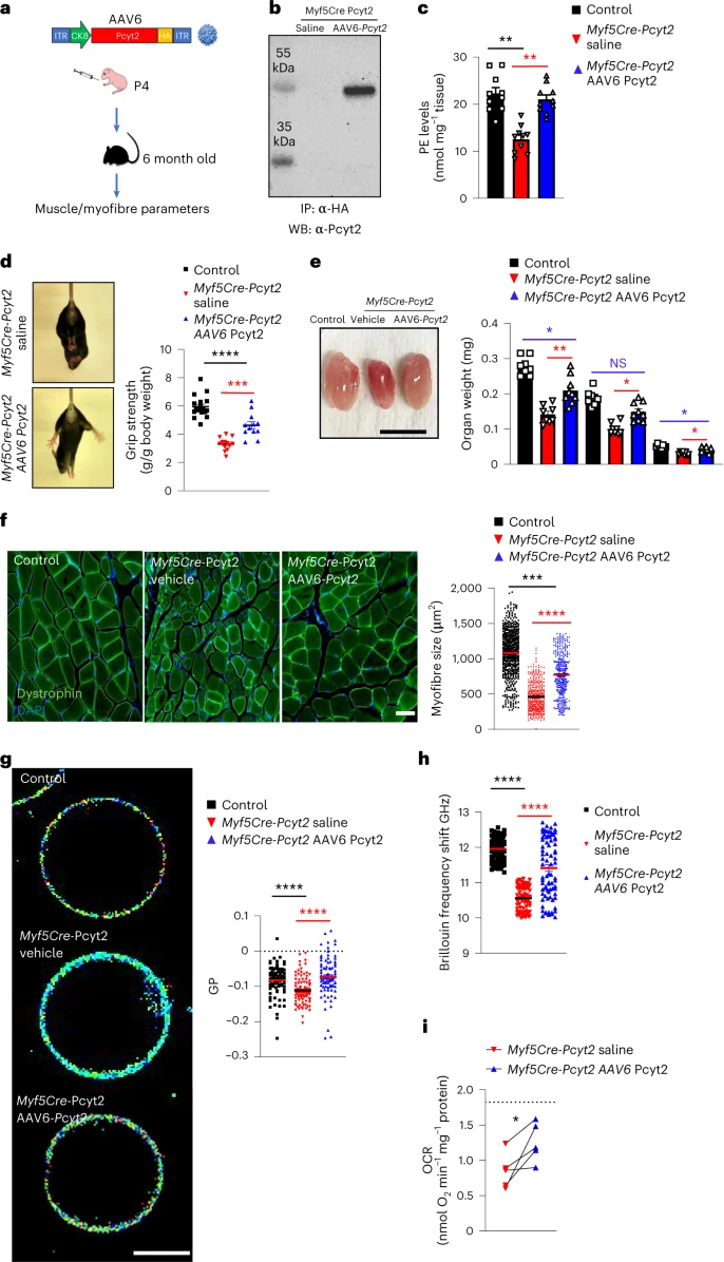
图7 腺病毒介导的Pcyt2基因治疗可有效治疗Pcyt2缺陷所致的肌肉病变
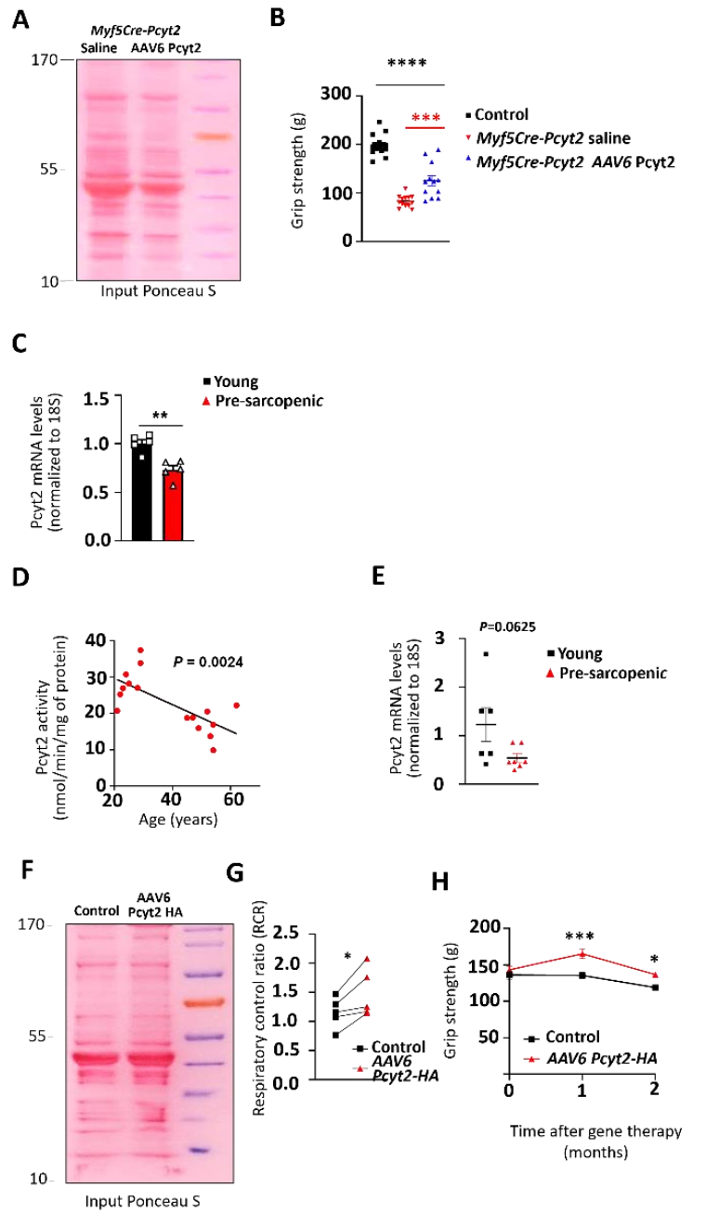
扩展图14 Pcyt2敲除小鼠和衰老小鼠的基因治疗效果
10 肌肉特异性Pcyt2基因治疗改善衰老中的肌肉健康
进行性肌肉萎缩是衰老虚弱的关键决定因素。肌肉衰老通常与膜完整性降低、损伤易感性增加以及运动后修复减少有关。由于Myf5-Cre-Pcyt2小鼠衰老后也出现肌肉退行性特征,作者评估了Pcyt2在肌肉衰老中的潜在作用。实际上,与年轻小鼠相比,在老年和患有肌肉减少症小鼠中四头肌中的Pcyt2 mRNA表达和酶活性降低(图8a,扩展图14c)。重要的是,与20至30岁的人相比,45至62岁的人的四头肌活检样品中,PCYT2活性和水平显著降低(图8b,扩展图14d-e)。因此,在小鼠和人类中,Pcyt2表达量和酶活会随着年龄的增长而下降。与此同时,作者发现,在衰老小鼠的肌肉中,受影响最大的脂质是各种PE脂质(图8c)。为了探究增加Pcyt2的水平是否可以改善老年小鼠的肌肉功能,作者利用上述AAV来处理小鼠,过表达Pcyt2后治疗衰老的肌肉。作者将改造的AAV 注射到24月龄雄性C57 B6/J小鼠眼眶后静脉窦中(小编注:腹腔注射和眼眶后注射用的病毒滴度不一样,腹腔注射是2×10^11,静脉注射是5×10^12)(图8d)。2个月后,四头肌中Pcyt2的表达和肌肉中PE水平显著增加(图8e-f,扩展图14f)。
此外,AAV的治疗效果也反映在肌纤维膜物理参数上,如肌膜硬度增加(图8g,h)。此外,肌肉的生物能量学显示出有益的变化,具有改善的线粒体能力和氧化呼吸能力(图8i,扩展图14g)。与此同时,在治疗2个月后,治疗组小鼠的肌纤维直径比对照小鼠更大(图8j)。值得注意的是,作者在该组小鼠中观察到治疗后1个月和2个月时握力改善(图8k,扩展图14h)。总之,以上数据说明肌肉特异性过表达Pcyt2可以改善肌肉衰老。
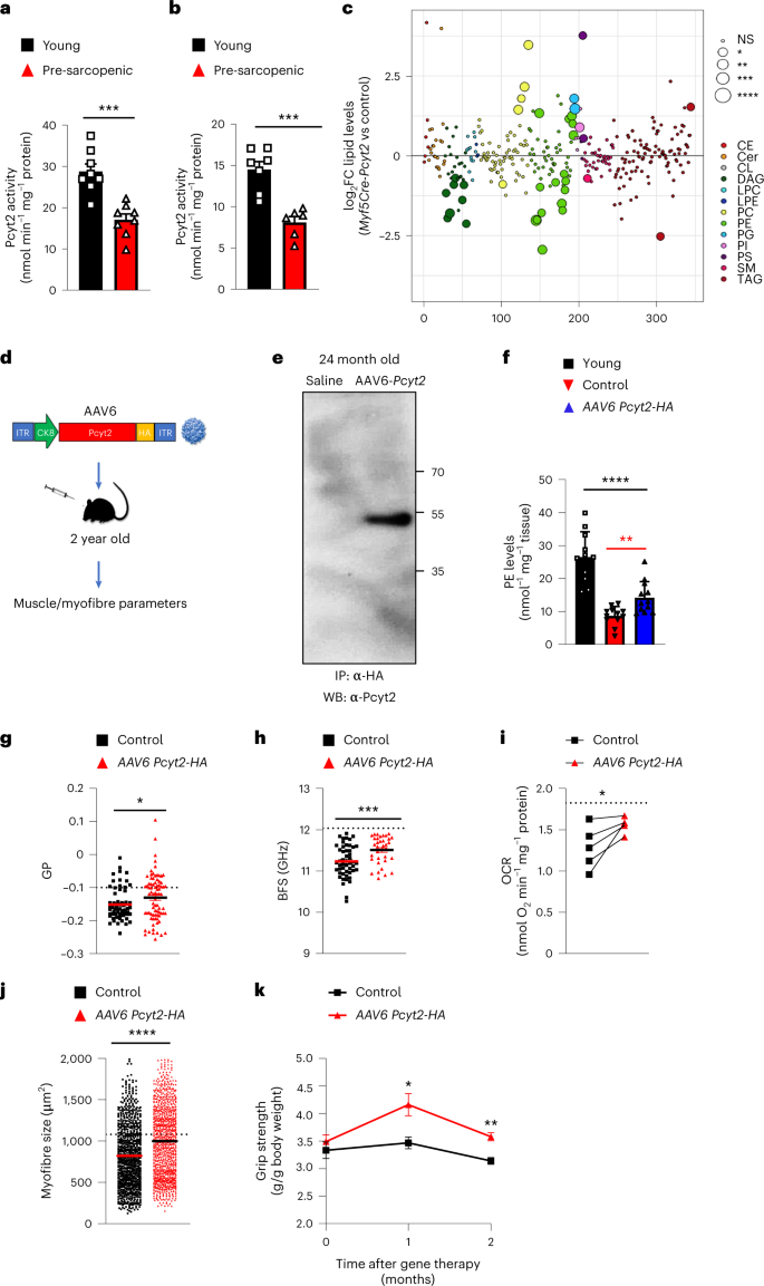
图8 人和小鼠衰老肌肉中Pcyt2活性降低,Pcyt2基因递送改善肌肉减少小鼠中年龄相关性萎缩
总结
肌肉退化普遍会在遗传性疾病或衰老中导致机体虚弱,阐明肌肉疾病的形成机制以及有效的治疗方法,是改善人类健康的重要目标。在本篇文章中,作者指出脂质合成酶磷脂酰乙醇胺胞苷转移酶(PCYT2/ECT)对肌肉健康是至关重要的。人体中的PCYT2缺陷则会导致严重的疾病,并伴随生长停滞和持续性虚弱。pcyt2突变的斑马鱼和肌肉特异性Pcyt2敲除小鼠表现出同样的表型,即生长停滞,进行性肌无力和衰老加速。在生理机制上,肌肉Pcyt2的缺乏影响细胞生物能量和膜脂质双层结构和稳定性。在在小鼠和人类中,肌肉衰老会导致PCYT2活性下降,并且基于腺相关病毒的PCYT2递送改善了Pcyt2敲除小鼠和衰老小鼠的肌无力,为患有罕见疾病和肌肉衰老的个体提供了治疗方法。因此,PCYT2在脊椎动物肌肉健康中起着极其重要的作用,将PCYT2和PCYT2合成的脂质与严重的肌萎缩/衰老联系起来。
原文链接:https://www.nature.com/articles/s42255-023-00766-2
https://m.sciencenet.cn/blog-3483272-1401313.html
上一篇:代谢学人--Cell Metabolism:看线粒体如何“嵴”速享“瘦”
下一篇:代谢学人--Cell Metabolism:储备粮草“脂质”与肿瘤增长的秘密