博文
代谢学人--代谢典藏 | Cell子刊综述:走向共识——哺乳动物脂肪干细胞和祖细胞的异质性
||
代谢典藏 | Trends in Cell Biology 综述
走向共识——哺乳动物脂肪干细胞和祖细胞的异质性

撰文 | 郭明伟 左慧 姚静 张俊
编辑 | 孟美瑶
引言
宋代文人苏轼在贬谪途中,与友人游观庐山时感慨道:横看成岭侧成峰,远近高低各不同。不识庐山真面目,只缘身在此山中。这首七言绝句形象地写出了庐山移步换形、千姿万态的风景。
咱们今天的主角“哺乳动物脂肪干细胞和祖细胞”也是如此。随着科学的不断发展,单细胞转录组学逐渐崭露头角,它为解析脂肪干细胞和祖细胞的异质性提供了一个全新视角。2020年10月23日,Cell系列综述期刊Trends Cell Biol.上在线发表的一篇题为"Toward a Consensus View of Mammalian Adipocyte Stem and Progenitor Cell Heterogeneity"的综述,接下来快跟随小编的步伐,探索脂肪干细胞和祖细胞的奥秘吧~

▼ 重点(Highlights)
1、脂肪干细胞和祖细胞(Adipose Stem And Progenitor Cell,ASPC)是异质性的,至少包括三个亚群:脂肪干细胞(Adipose Stem Cells,ASCs)、前脂肪细胞(Preadipocytes,PreAs)和脂肪生成调节因子(Adipogenesis Regulators,Aregs)。
2、PreAs(ICAM1+,VAP-1+)和Aregs(CD142+)均起源于ASCs(DPP4+,CD55+)。
3、在成年小鼠中,Aregs(CD142+)通过旁分泌机制抑制脂肪生成,但具体机制尚不完全清楚。
4、在内脏脂肪中,ASCs的一个亚群——纤维炎性祖细胞(Fibro Inflammatory Progenitors,FIPs)(PDGFRβ+/LY6C+)具有抑制脂肪生成的作用。
5、间皮细胞(Mesothelial cells)分布于内脏脂肪的血管基质组分(SVF),能够调节免疫微环境,但并不分布于皮下脂肪组织中。
6、人和小鼠来源的ASCs表现出共同的基因表达特征,但人的PreAs和Aregs在分子上的差异不如在小鼠中明显。
7、由于人和小鼠的ASPC之间存在异质性,这种差异引发了对“小鼠ASPC是合适的人类脂肪生物学模型系统”这一结论是否完全正确的质疑。
摘要
白色脂肪组织(White adipose tissue,WAT)是一种细胞异质性的内分泌器官,不仅充当能量贮存库,而且积极参与代谢稳态调控。脂肪组织的主要成分是成熟脂肪细胞,其起源于脂肪干细胞和祖细胞(ASPCs)。众所周知,这些ASPCs存在于脂肪组织的血管基质组分(SVF)中,但尚不清楚它们的分子异质性和功能多样性。近年来,随着单细胞转录组学解析能力的提高,最近的几项研究为不同种类哺乳动物脂肪组织中ASPCs的细胞异质性提供了新的见解。本文就近年来有关ASPC的研究进展、它们的细胞亚群结构、层次结构、各脂肪库独特性质、功能和调控机制等方面作一总结,并讨论了小鼠ASPC与人类ASPC生物学特性的异同。
脂肪干细胞和祖细胞:成熟脂肪细胞的前身
WAT是以脂肪细胞为基本单位的疏松结缔组织,存在于多个解剖学位置(如皮下脂肪和内脏脂肪)。脂肪细胞周围的结缔组织含有血管基质组分(SVF),其中存在脂肪干细胞和祖细胞(ASPCs),是成熟脂肪细胞的前身。它们与免疫细胞和脉管系统一起,维持整个组织的稳态。
ASPCs被定义为不含免疫细胞(CD45+)、内皮细胞(CD31+)和红细胞(TER119+, CD235a+)的SVF细胞,也称SVF的谱系阴性组分(Lineage negative fraction,Lin-)。它本身是一种异质性细胞群。因此,解析ASPCs中的细胞谱系层次结构是该领域的一个长期挑战。谱系研究的基本观点认为,一定有一个ASPCs亚群是脂肪细胞生成的根源,即“真正”的脂肪干细胞,而其他ASPCs则代表了向成熟脂肪细胞定向分化过程中的各种瞬时状态。为了进一步验证这一假说,我们需要了解ASPC的细胞异质性究竟如何,以及是否存在不同的分子和免疫表型的ASPC细胞亚型。另一个重要的问题是内脏脂肪组织和皮下脂肪组织的ASPC细胞图谱有何不同,这些差异在多大程度上反映了不同脂肪组织的生理特性。最后,考虑到肥胖中脂肪细胞增生(Hyperplasia)相对于脂肪细胞肥大(Hypertrophy)更具有益处,以及肥胖中内脏脂肪扩张相对于皮下脂肪扩张更具风险,在脂肪过度积累的大背景下解析ASPC异质性别具意义。
在小鼠模型谱系追踪(Lineage tracing)方法的基础上,单细胞RNA测序或单细胞转录组学(scRNA-seq)的发展为识别和表征ASPCs并回答上述问题提供了前所未有的机会。如后文将要讨论的,谱系追踪一次只能探索一个或几个基因标记,因此谱系追踪方法的主要缺陷是研究范围有限。而全基因组检测(如scRNA-seq)能为特定生物系统(如ASPCs)的异质性提供全新的视角。但是scRNA-seq并不是一种简单的实验室技术,特别是在分析由此产生的大数据集时尤为困难。因此,目前已经开发出在线平台来共享scRNA-seq分析。
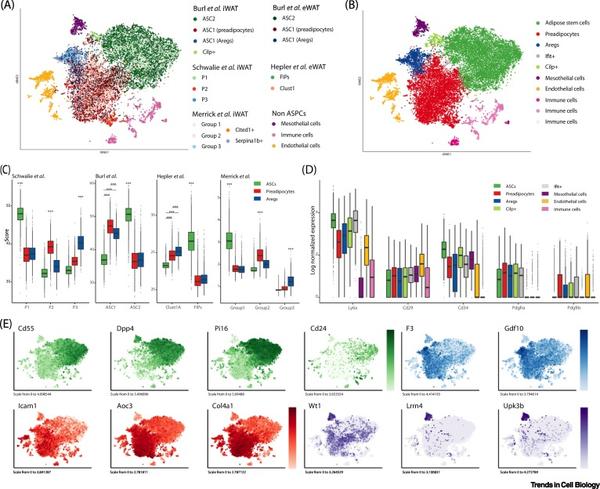
2018年, scRNA-seq被首次用来解析小鼠脂肪SVF异质性,随后又有了针对其他几种脂肪的scRNA-seq分析。在本综述中,我们将讨论这些研究如何极大地推进了我们对哺乳动物WAT中ASPC的组成和功能的理解。具体来说,ASPCs的scRNA-seq研究确定了3-4个细胞亚群,这些细胞亚群间具有沿分化成脂轨迹排序的层次结构关系。为了比较这些不同研究的结论,作者对各种scRNA-seq数据集进行了全新的整合分析(图1和图S1)。为了更好地说明和比较不同的细胞亚群,作者基于分析结果及报道的ASPC各亚群的分子和功能特征,提出了一种广义的命名术语(Coarse-grainconsensus nomenclature):脂肪干细胞(Adipose stem cells, ASCs)、前脂肪细胞(Preadipocytes, PreAs)、脂肪生成调节因子(Adipogenesis Regulators, Aregs)。在下文中将根据每个亚群在层次模型中的位置,逐一着重介绍。
ASPC异质性
在小鼠中,由Ly6a基因编码的SCA1膜表面蛋白是一种广泛认可的可用来富集具有体外成脂分化能力的前体细胞群的ASPC主要标志物。然而,并非所有的SCA1+细胞都能在体外产生脂肪细胞,提示可能存在更精细维度的细胞状态或细胞类型。因此,研究人员投入了大量的精力来寻找更多的表面分子标记物。例如,间充质标记物,包括CD34、CD29、CD24和血小板来源的生长因子受体 (PDGFR)α/β,这些分子标记被证明富集于成脂前体细胞,或其中的一部分。作者的数据整合分析证明几乎所有ASPC都表达Cd34、Cd29和Pdgfra,而Cd24和Pdgfrb的表达则不具有广谱性(图1D,E)。虽然SCA1在小鼠中被广泛用于富集ASPCs,但在人类中却没有SCA1的同源基因。因此, SVF中的整个谱系阴性(Lin-)组分被认为最好地代表了人源ASPCs(hASPCs)。CD29、CD34、CD13、CD44、CD73、CD90、CD142、CD9、CD10和CD200等标记物已被用于进一步富集hASPCs,然而对于hASPCs的精确分子特征尚未达成共识。目前,这些标记物的鉴定大多基于流式细胞术和免疫组化技术,并以造血、内皮和神经组织为参照物,因此会造成标记物的选择偏差。然而,在过去的几年里,单细胞技术的革命性发展使得研究人员能够以前所未有的分辨率解析ASPC(图1和图2)。
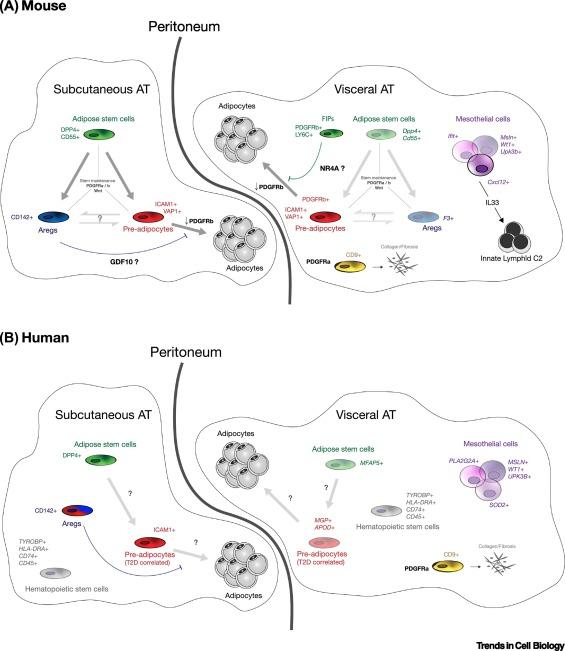
缩写:AT,脂肪组织;IL,白介素;ICAM1,细胞间粘附分子1;PDGFR,血小板源性生长因子受体;T2D:2型糖尿病;VAP1血管粘附蛋白1
白色脂肪组织的三个主要亚群
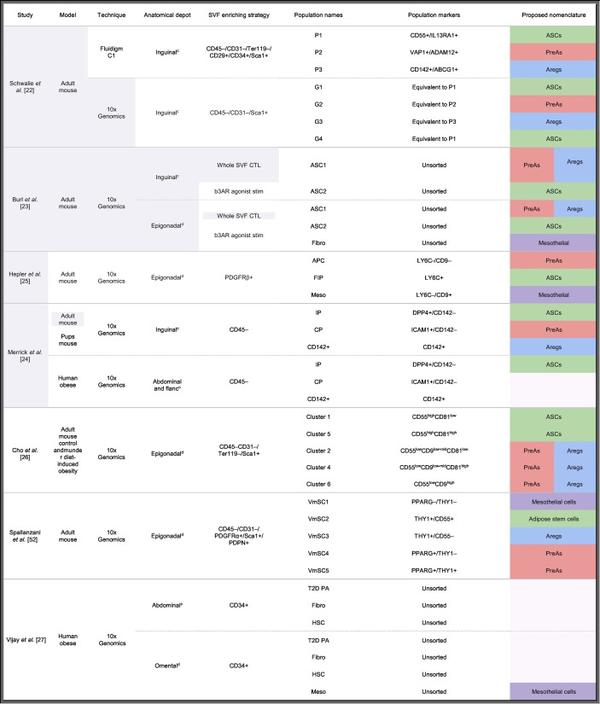
近年来许多研究解析了小鼠皮下(Inguinal WAT,iWAT)和内脏(尤其是性腺周围脂肪,Epigonadal WAT,eWAT)白色脂肪中ASPC的异质性。尽管所使用的方法和小鼠模型不尽相同(表1),但所有研究都将ASPCs区分为两到三个主要亚群。细胞亚群特异性的分子标记物定性交叉比较(Qualitativecross-comparison)证明皮下和内脏脂肪中都存在这些细胞亚群,尽管它们的内在差异仍未完全了解。为了对ASPC分类进行验证,作者使用Seurat标准工作流程整合分析了公共数据库数据(表1中以灰色突出显示)。通过对这个庞大的集成数据集进行深入研究,可以无偏向性对各研究中所鉴定的ASPC图谱进行探索。这是因为它可以验证先前鉴定到的ASPC表面标记分子的表达谱和表达特异性。此外,通过这些数据集作者可以探索近年来发表的scRNA-seq研究之间的相似之处,并支持提出的细胞亚群命名术语。具体来说,作者的分析鉴定结果强烈支持存在三个细胞群体(ASCs、PreAs和Aregs),主要的理由是:(1)各自具有特定的基因表达特征;(2)首次在每个数据库中独立检测到;(3)在不同的数据集中可以相互验证(图1A,图S1A、G);(4) 通过对整合数据集的聚类分析,一致地被分别聚类到各个细胞亚群中(图1B,图S1B、H)。最后,作者在本综述中鉴定的三个亚群与之前不同研究发现的亚群一致(图1C和图S1C、D、I、J)。总之,整合分析表明与PreAs和Aregs相比,ASCs是一个不同的细胞亚群,PreAs和Aregs在基因表达方面更接近,但仍具有不同的转录谱。除此之外,间皮细胞也与ASPC一起构成内脏SVF的Lin-组分,但它们被认为与ASPC不属于一个层次结构,因此将在框1中单独讨论。
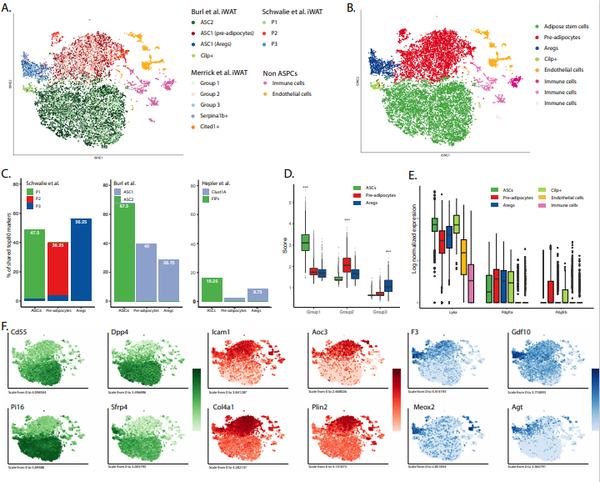

F,L:tSNE图展示了选定标记(Aoc3,VAP1;和F3,CD142)的对数归一化表达,标记不同颜色。颜色反映亚群特异性(绿色,脂肪细胞干细胞(ASC);红色,前脂肪细胞(PreAs);蓝色,脂肪生成调节细胞(Aregs);紫色,间皮细胞)。
d.Visceral(内脏)。
▼ 框 1:间皮细胞
间皮细胞(Mesothelial cells)构成了小鼠和人的腹膜,是内脏SVF的特异组分。它们不仅构成腹腔壁,在visWAT内部也成簇存在,也即所谓的“乳白色斑点”或“脂肪相关淋巴团簇”。Msln、Wt1、Lrn4和Upk3b是间皮细胞的典型分子标记物,在体外,这些细胞显示出典型的上皮/鹅卵石形态。通过scRNA-seq在内脏ASPCs中也发现了间皮细胞(鼠[23,25,28],人[27],图1A、B、E,图S1G、H、I)。因此,必须注意的是,在人类和小鼠的内脏SVF中都有间皮细胞存在,因此可能干扰功能试验结果。作者认为在进行ASPCS的任何下游体外实验验证前,最好先将间皮细胞组分去除后。
然而,在脂肪生物学的背景下,间皮细胞的有趣之处在于,除了其本身是一种异质性群体外,它们在脂肪功能和免疫间的桥梁作用,以及其作为肿瘤转移热点的角色。当腹膜炎发生时,它们招募中性粒细胞以清除腹膜液,而在生理条件下,它们支持白细胞介素(IL)-33驱动的2型先天淋巴样细胞(ILC2s)招募和分化。ILC2细胞分泌2型免疫调节细胞因子,与调节性T细胞和嗜酸性粒细胞共同作用,确保内脏腔和脂肪组织保持在正常、无炎症状态,并抵抗2型糖尿病的发展。相反,IL-33的丢失会导致体重增加,尽管其机制、以及ASPCs在这一过程中的潜在作用机制尚不清楚。对于间皮细胞免疫调节作用的更详细介绍,请见本综述参考文献。
在人类中,间皮细胞、免疫细胞和基质细胞之间的相互作用知之甚少。然而,有研究提出间皮细胞亚群与米色脂肪形成有关,因为这些亚群的线粒体基因表达比例高于任何其它ASPC亚群,并且其相对细胞数量与空腹血糖水平呈负相关。此外,米色脂肪细胞分子标记物PLA2G2A和SOD2在间皮细胞亚群中富集,而两种棕色脂肪前体细胞标记物MYF5和ZIC1没有富集,为这些人间皮细胞参与米色脂肪形成提供了额外的支持和证明。
对人类研究而言,目前只发表了少数scRNA-seq研究,而且这些研究间的结论并不一致,这使得确定hASPC的异质性很难。然而对已发表结果的仔细分析可发现,基于共同的基因特征,至少有两个ASPC亚群在小鼠和人类之间是保守的,即ASCs和PreAs。
脂肪干细胞群
使用scRNA-seq发现的第一个ASPCs亚群([22]中的P1,[24]中的Group1,[23]中的ASC2)在本质上最像干细胞,因此作者提议将这一亚群称为ASCs(图1A-C)。ASCs不仅表达干细胞相关基因,如Ly6a、Dpp4、Cd55和Wnt信号通路抑制因子Sfrp4,还缺乏某些其他基因(如编码 CD142的F3)(图1D,E),而且在体外具有高度增殖能力。从功能角度来看,当单独用胰岛素进行体外诱导时,ASCs[CD142-/DPP4+(二肽基肽酶4)]往往表现为成脂抑制,且不表达脂肪细胞特异基因。但是使用完全成脂分化液(Complete adipogenic cocktail,CAC)诱导时,ASCs (CD142-/DPP4+或SCA1+/CD55+)表现出相对较高的成脂分化潜能,但其分化程度在不同研究中存在差异。这些结果表明ASCs需要关键因子来决定其细胞命运和分化潜能,从而产生具有不同成脂潜能的细胞状态。因此,ASCs很可能是一种间充质干细胞,只有当暴露在正确因子下(如CAC)才会形成脂肪细胞。此外,它们也能被诱导形成成骨细胞,并表达成骨细胞特异性标志物(Alpl、Bsp、Osx和Ocn),这表明了其多能性。
总之,在两项不同的研究中,Lin-/SCA1+/CD55+和Lin-/CD142-/DPP4+细胞代表了两种相似的、具有类似干细胞特征的分子和功能特性的细胞亚群。到目前为止,ASCs为何具有干性的分子机制仍不清楚。转化生长因子(TGF)β是一个脂肪抑制因子,它能够调节DPP4+细胞群的命运和功能。具体而言,TGF β处理能够提高ASCs的增殖能力,抑制脂肪细胞形成(即使在使用CAC的情况下),并上调ASCs特异性分子标记表达,而抑制TGFβ信号通路则有相反的作用。值得注意的是,TGFβ激动剂和拮抗剂对其他ASPC亚群均无影响,这也凸显了TGFβ信号通路对DPP4+细胞群的特异作用。抑制成脂的Wnt信号通路在调节新生脂肪生成中似乎也有着重要作用,因为当β-catenin水平超过TCF7L2时,细胞倾向于维持其干细胞状态,而反之则诱导脂肪生成。总的来说,这些研究表明多种信号通路参与维持ASC,但确切的分子机制仍不清楚,包括这些通路在多大程度上会相互影响?什么因子触发了这些信号通路?以及它们如何影响整个细胞状态?
在人类研究中,尽管第一个较低分辨率的单细胞测序研究报道了hASPCs之间具有“同质性”,但该研究也得出与小鼠研究类似的结论,如CD55和THY1等基因在各个细胞中并不是均一表达的。的确,之后的研究发现了一个表达DPP4、CD55和MFAP5的hASPC亚群,其转录谱特征类似小鼠ASCs。另外,人源ASCs只有在受到几种成脂诱导因子的组合刺激下才显示出完全的成脂潜能,单独使用胰岛素无法诱导其成脂,这点也与小鼠相似。有趣的是,表达THY1的间充质干细胞被认为是导致2型糖尿病患者肌肉脂肪变性的原因,这个亚群是否在转录谱上与hASCs相似还有待证实。
内脏脂肪中的ASCs
在小鼠eWAT的scRNA-seq数据集中发现了一个具有类似ASCs分子和功能特性的群体([23]中的ASC2和[26]中的Clusters 1和5,图1B-E,图S1G-J)。例如,与其他内脏APSCs相比,内脏ASC(Lin-/SCA1+/CD55HIGH细胞)的增殖能力更高,成脂分化能力更低。此外,内脏ASC(Lin-/CD142-/DPP4+细胞)表现出较低的成脂分化潜能,对TGFβ处理的反应性则与皮下ASC类似。有趣的是,eWAT中ASC的相对丰度低于iWAT,表明内脏WAT (visWAT)具有更小的早期干细胞群。
在机制上,PDGF通路在脂肪分化和ASC干性维持中起着关键作用,这方面的研究主要集中在eWAT中。PDGF受体与干细胞维持有关,需要下调PDGF受体来启动脂肪生成。研究表明,主要脂肪库中的成熟脂肪细胞均来自Pdgfra+细胞,且PDGFRα的表达先于PDGFRβ。也有研究报道 PDGFRβ+脂肪前体细胞能够响应高脂饮食(HFD)分化为脂肪细胞。根据作者对scRNA-seq数据集的分析,Pdgfrα在所有ASPC中几乎都是阳性的,而Pdgfrβ则只在ASC中以非常低的水平表达(与ASPC群的其余部分相比)(图1D,图S1E,K)。这些结果支持了这样一种观点,即ASC是“真正的”脂肪干细胞,能够产生命运更确定的祖细胞,进而分化为脂肪细胞。
PDGF通路在细胞干性和脂肪生成中的重要作用促使研究者利用scRNA-seq特异性地表征PDGFRβ+祖细胞群。通过该方法鉴定出了一个与ASCs存在许多相同标志物的PDGFRβ+细胞群(图S1l)。这群PDGFRβ+/LY6C+细胞群被称为纤维炎性祖细胞(FIPs)。与之前在ASCs中的发现一致,对FIPs基因表达谱的分析显示出存在活跃的TGFβ信号通路,而TGFβ处理能进一步上调FIP特异性标志基因,尤其是胶原相关基因(命名中“纤维”二字的由来)。同时,FIPs也表现出一种功能性促炎表型,表达多种细胞因子如Il6、Cxcl2和Cxcl10,并显示出在体外激活巨噬细胞的能力(命名中“炎性”二字的由来)。最后,FIPs成脂能力低且取决于培养条件,提示其与ASCs类似。然而,研究者也揭示了FlPs能够通过一种尚未明确的分泌因子发挥抑制成脂的功能。因此,即使内脏FIPs与ASC有共同的重要分子标记物(图1A、C,图S1l),它们与皮下ASC又具有明显不同的表型特征,因此很难建立明确的类比关系。的确,由于皮下脂肪中所有PDGFRβ+细胞都是LY6C+,所以皮下脂肪中的ASPCs不能用基于FIPs研究的分类标记物进行区分。因此,目前FIPs算是一个略显神秘的独特细胞亚群,它可能反映了(内脏)ASC响应特定的免疫刺激或生理条件而改变自身细胞状态,发挥功能的一种内在能力。
前脂肪细胞群
通过scRNA-seq发现的第二大也是最丰富的ASPC细胞亚群,它们以表达Icam1和Aoc3以及几种胶原蛋白和细胞外基质重构因子为特征,作者在这里将其定义为PreAs(图1E和图S1F)。到目前为止,还没有研究专门聚焦PreAs。然而, PreAs的分子特征以及初始功能特征都标明,其与ASCs相比具有更确定的成脂细胞命运(Committed Adipogenic State)。首先,多个参与脂肪生成的基因,如Pparg、Fabp4、Lpl、Plin2或Cd36,都在这个细胞亚群中显著上升([22]中的P2,[24]中的Group 2,[23]中的ASC1;图1E,图S1F);其次,与ASC相比,PreAs的增殖能力较低,而成脂能力较高,仅由胰岛素刺激便可表现出高成脂分化潜能。总的来说,Lin-/CD142-/ICAM1+(细胞间黏附分子1)细胞亚群和Lin-/SCA1+/VAP1+(血管黏附蛋白1)细胞亚群似乎是具有成脂命运的相似细胞亚群。在机制上,PreAs 对TGFβ的成脂抑制效应最为抵抗,进一步支持PreAs是具有成脂谱系命运的ASPCs。
在人体研究中,也同样鉴定出了类似于小鼠PreAs的细胞亚群,其表达ICAM1、PPARγ和GGT5。在体外实验中,人类PreAs的表现与小鼠类似。
Burl等人在内脏和皮下脂肪细胞中鉴定出一个细胞亚群 ([23]中的ASC1),该细胞群与PreA细胞群类似(图1B-E,图S1G-J)。作者对该研究的数据集进行分析发现,ASC1亚群具有与PreAs和Aregs类似的高表达基因标志(CD142+),因此可以进一步对其进行分群(图1C,图S1C、I)。尽管该研究主要是计算性研究,但其后一项平行研究发现了一群由Lin-/PDGFRβ+/LY6C-/CD9-定义的细胞群,基于作者的数据整合分析,该细胞亚群部分投射到PreAs(和Aregs)(图1A、C,图S1G、H)。分离出的Lin-/PDGFRβ+/LY6C-/CD9-细胞亚群与Lin-/PDGFRβ+细胞群相比,具有更高的成脂分化潜能,这与皮下PreAs的特征一致。因此,该研究的作者可能从visWAT中分离出了一群PDGFRβ+ PreAs亚群。另一项独立研究则发现了三个具有更高成脂分化能力的亚群 ([26]中的Cluster 2、Cluster 4和Cluster 6),这三个细胞亚群都部分投射到PreAs和Aregs上。
脂肪生成调节因子亚群
iWAT中的Aregs
特征性表达F3(编码CD142)的第三个ASPC细胞亚群在成年小鼠的iWAT中被首次报道,与ASCs相比,其基因表达谱更接近PreAs(图1C、E)。在功能上,这些细胞不仅在体外抵抗成脂分化,而且在体外和体内实验中,还具有抑制其他ASPCs组分成脂分化的能力。因此,作者将这群细胞命名为Aregs(图S1F)。
由于其独特的转录组聚类模式,第二项研究也关注到了CD142+细胞亚群,但与第一项研究相反,却发现CD142+细胞亚群具有完善的成脂分化能力。虽然这两项研究在方法和小鼠模型上有着明显差异,但CD142+ ASPCs在不同研究中为何表现如此不同,原因仍待探究。不管如何,目前的共识是,CD142+ ASPCs构成了一个独特的细胞亚群,其具有明显不同于ASCs和PreAs的转录组特征(图1B、C,图S1C、F)。
除了在脂肪组织中,最近已经在肌肉中发现了类似Areg的细胞,具有与皮下脂肪Aregs类似的分子和功能(例如抑制脂肪形成)特性。作者提出了一个机制模型,认为在小鼠和人中CD142+细胞亚群主要通过分泌因子GDF10起到抑制成脂的作用。有趣的是,研究发现,肌肉营养不良小鼠中的“肌肉Aregs”比野生型(WT)小鼠要少,提示了这一细胞组成上的变化可能是萎缩的肌肉中脂肪细胞过度累积的原因。
对于人类中的Aregs细胞亚群,发现小鼠Aregs分子标记物(CD142、CLEC11A和FMO2)在人的两个主要细胞亚群ASCs和PreAs中广泛表达。因此,作者认为,可能不存在和小鼠同源的人类Aregs细胞亚群,也可能其在转录组水平上不能被小鼠的分子标记物所定义。与此一致的是,分选出的人源CD142+细胞和ICAM1+ ASPC之间没有发现功能差异。然而,当对人源CD142+细胞进行分离并与相应CD142-和Lin-组分进行比较时,也在人源CD142+ ASPCs中观察到了与小鼠Aregs类似的非成脂特性。因此,未来需要对人源CD142+细胞亚群的分子和功能特征研究进行更深入的解析。
visWAT中的Aregs
目前尚无针对visWAT的scRNA-seq研究专门探讨Aregs的存在,另外,尽管FIPs和Aregs均表现出成脂抑制的表型,二者在分子特征上明显不同(图1C)。尽管使用流式分析发现与iWAT相比,eWAT中CD142+细胞的比例更高,但目前还没有研究对内脏脂肪Aregs进行完善的分类和鉴定。然而,最近一项研究发现了一个ASPC亚群(Lin–/Sca1+/PDGFRA+/PDPN+/THY1+/CD55–细胞),其转录组特征与Aregs相似,成脂分化能力低,虽然这群细胞并不是使用CD142表面分子标记物分离得来的。与此一致的是,作者的整合分析结果也在eWAT中发现了一种表达Areg样分子标记物的细胞亚群(图S1H、I)。令人惊讶的是,尽管F3是此细胞亚群的差异表达基因(DE)之一,但缺乏特异性,因此不是该亚群的最佳分子标志物(图S1F与图S1L)。F3这种广泛的、非特异的表达模式可能与visWAT流式分析中观察到的较高的CD142+比例有关,并不一定表明其中具有高比例的Aregs。但是,使用从scRNA-seq数据推断出的相对细胞比例,同时考虑到这种方法会由于细胞选择而产生可能的数据偏倚,在内脏脂肪数据集中表达Areg样标志物的细胞比例依然显著高于皮下脂肪(仅考虑ASPCs,eWAT为23.5%,iWAT为7%)。内脏脂肪中具有更高比例的抑制成脂性的ASPC(可能同时包括“内脏Aregs”和FIP两种细胞),可能与“内脏ASPCs比皮下脂肪ASPCs具有更低的成脂分化潜能”这一领域共识相一致。这些发现不仅揭示了不同脂肪组织库中内在的相似性,更强调了其中的差异,说明各个ASPC组分的功能值得更深入地研究。
ASPCs的层次结构
根据本综述中对不同ASPC亚群(ASCs、PreAs和Aregs)的命名,作者提出ASPC之间的层次结构(图2中箭头,关键图)。首先,如前文所述,Pdgfra和Pdgfrb的表达模式表明ASCs是最类似干细胞的ASPCs。Cd24在ASC中的高表达进一步支持了这一假设(图1E)。同时有研究发现CD24+脂肪细胞前体是干细胞样ASPC,它们通过失去CD24表达而进一步获得脂肪细胞的命运。这一层次结构也被近期的在scRNA-seq数据集上应用拟时序轨迹分析的研究所揭示,并进一步使用基于荧光的细胞追踪技术在体内进行了验证。具体而言,将荧光标记的皮下ASC(DPP4+细胞)移植到WT皮下脂肪垫中时,这些细胞在移植后7天内表达PreAs和Aregs的分子标记物(分别为ICAM1和CD142),并且其中一部分细胞在14天后不再表达DPP4。这些研究表明PreAs(ICAM1+)和Aregs(CD142+)均由ASC(DPP4+)产生而来。相比之下,荧光标记的PreAs(ICAM1+)和Aregs(CD142+)不会随时间延长而获得DPP4表达,表明了ASC命运确定和分化的整体单向性。然而,PreAs和Aregs可能会发生免疫表型互换,例如,发现一部分mTomato标记的ICAM1+和CD142+细胞在一定程度上分别获得了CD142和ICAM1标记。这种互变现象在分子上是如何进行编码的,是什么决定了细胞亚群之间的平衡,以及不同的代谢条件是如何影响这种平衡,这些是重要而尚未解决的问题(框2)。
▼ 框 2:ASPCs与肥胖
ASPCs与肥胖
目前,我们对ASPC在肥胖中的动态变化知之甚少,各个亚群对于肥胖的影响则了解更少。目前已知的有,在瘦小鼠和饮食诱导的肥胖小鼠的eWAT上进行了scRNA-seq测序,揭示了肥胖会影响ASPC各亚群的相对数量。此外,肥胖特异性增加了具有较强细胞外基质分泌和免疫调节能力的ASPC亚群的丰度,并改变其分化能力。与此一致的是,当给小鼠喂食HFD时,FIPs数量增加,而DPP4+细胞比例下降。总之,这些研究表明ASCs在WAT中具有重要的免疫调节作用,这与之前的研究结果一致。同时说明饮食诱导的肥胖中ASC向FIP样表型的偏移失衡是肥胖引起的脂肪炎症的潜在因素之一。
PreAs与肥胖
有假设认为ASPCs的内在特性可以解释在肥胖期间不同解剖位置的脂肪沉积对代谢性疾病发展的独特影响(另见综述)。与此相符的是,从肥胖小鼠或瘦小鼠中分离的皮下脂肪ICAM1+细胞都具有相对较高的成脂能力。而与瘦小鼠相比,肥胖小鼠的内脏脂肪ICAM+细胞成脂分化潜能更低。该结果可以解释为什么腹部脂肪更倾向于肥大性WAT扩张而非增生性WAT扩张。
有趣的是,根据来自25个肥胖患者的大型scRNA-seq数据集,人类PreAs比例的相对增加似乎与胰岛素抵抗和2型糖尿病相关。据我们所知,这是在人类中首次发现代谢疾病表型与ASPC组成差异之间具有相关性。这种仅基于scRNA-seq细胞比例的相关性是否能反映因果关系仍有待确定。
Aregs与肥胖
WAT中可能含有能够负向调节脂肪细胞形成的ASPCs,如Aregs或FIPs,这一事实刷新了我们对脂肪组织可塑性的认知,提出了潜在的新型治疗应用。例如,抑制脂肪生成的细胞比例减少可能是导致脂肪细胞增生的机制之一。相反,这些成脂抑制细胞的增多可能通过抑制新生脂肪细胞生成而导致脂肪细胞肥大。有趣的是,有研究表明,在高脂诱导的情况下,ASPC的耗竭是导致脂肪细胞由增生向肥大转变的原因,CD142+细胞的丰度在各个脂肪库和生理状态之间的变化也进一步支持这一概念。确实,尽管肥胖小鼠中CD142+细胞的增加看似不合理,但鉴于它们抑制脂肪形成的功能,这可能代表了机体为限制脂肪组织(过度)扩张而采取的一种防御策略。但这一机制的主要缺点是,在体重过度增加的情况下会更偏向于脂肪肥大而非增生。
hASPCs的特点
通过从25个肥胖个体的内脏和皮下脂肪的hASPC测序,得到了一个庞大的scRNA-seq数据集,其描述了在一些在转录组特征上与小鼠ASPCs相似的hASPC群体。因为这些细胞表达TYROBP、HLA-DRA和CD74等标志物,研究者将该群体的细胞定义为造血干细胞(HSC),然而至今尚未对这些细胞进行功能表征,也不清楚其生物学性质。值得注意的是,从技术角度来看,该研究在scRNA-seq分析前未对SVF进行CD45-分选(仅就CD34+进行了分选)。因此,这些HSC实际上可能是CD34+/CD45+细胞,应被视为“免疫细胞”而不是hASPC。无论如何,CD75被认为有助于巨噬细胞在肥胖环境中的促炎转换,并与胰岛素抵抗有关,故而在肥胖的情况下研究“HSCs”可能是有意义的。
尽管不是来自scRNA-seq,另外有其他几种表面标志物也显示出功能相关性,尤其是在人类中的。例如在小鼠和人类中,CD9标记了一组具有促纤维化特性的祖细胞亚群。而PDGFRA被报道介导ASPC中CD9low表达细胞向CD9high高表达细胞的转换,从而引发纤维化、炎症和胰岛素抵抗。CD9表达与上述ASPC群体存在多大程度上的重叠还有待研究。最后,由于迄今为止所有人类相关研究都在肥胖个体中进行,因此瘦人群中的hASPC组成暂未被解析,这是一个重要的未解决的科学问题。
总结与未来展望
近年来发表的多项研究使我们对ASPC的异质性有了更深入的了解,于此同时需要通过处理更多的细胞和总结现有的数据集来进一步揭示细胞分群的精细程度。因此,作者鼓励整个单细胞领域在各自的论文中始终公开其(sc)RNA-seq数据,这将更好地促进数据集整合,进而激发更大的研究兴趣。
已发表的研究报道解析了ASPC细胞群体的异质性,而对成熟脂肪细胞的异质性了解更少,但也不是毫无进展。例如,最近对来自单个祖细胞脂肪细胞克隆进行的批量转录组分析表明,成熟脂肪细胞具有异质性,但是最终仍需要对原代分离的成熟脂肪细胞进行深入的scRNA-seq分析,最终获得无偏见的脂肪细胞异质性图谱。探索成熟脂肪细胞在技术上比探索ASPCs更困难,脂肪细胞的脆弱性目前已经催生了利用细胞核转录组测序来替代细胞整体测序。初步结果提示脂肪细胞在分子和功能上存在广泛的异质性。该领域的终极挑战是将来自ASPC的scRNA-seq数据与成熟脂肪细胞的数据联系起来,在不同脂肪组织中清楚地建立起细胞谱系图谱。
另一个巨大的挑战是阐明人类与小鼠的脂肪组织中细胞组成在何种程度上相似。现有数据清楚的表明尽管各种细胞亚群有所重叠,人类和小鼠间仍存在巨大的差异,如二者间缺少保守的关键间质细胞标志物(如小鼠SCA1),而且与小鼠不同的是,人体内CD142+细胞亚群的分子性质尚不清楚。这表明除了代谢、行为和生理上的差异之外,细胞亚群异质性上的差异也会限制将小鼠肥胖相关的研究结果外推到人类中。尽管动物模型对于开发新的肥胖治疗策略仍是至关重要的,但要充分考虑其优势和局限性,仍需新的实验系统来专门研究人类脂肪生物学。一种有潜力的方法是利用类器官进行研究。近年来,越来越复杂的类器官模型已经可以模拟人体组织的关键结构和功能特性。因此,类器官模型可能具有巨大的生物医学影响,为直接干预人类细胞,并研究这些干预如何影响发育和功能提供了良好的平台。在下一阶段,甚至可以设想出更复杂的微生理系统,将其与其他器官(如肝脏)的类器官模型整合后,最终可以使我们在体外模拟肥胖的系统生理效应。
▼ 名词解释
原文链接:https://www.sciencedirect.com/science/article/pii/S0962892420301860?via%3Dihub
关注微信公众号代谢学人
了解更多代谢前沿资讯

https://m.sciencenet.cn/blog-3483272-1310929.html
上一篇:代谢学人--Science子刊:肌肉基因突变效果男女大不同
下一篇:代谢学人--Nature Metabolism 十月刊 饮食与营养精选